
Population Structure of Three Provenances of Vitis tiliifolia (Humb. & Bonpl. ex Schult.) in the Central Zone of the State of Veracruz, México

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
- Growth and formation period (seedling), herbaceous shoots of no more than 60 cm [19].
- Plant development period (juvenile), individuals with buds and shoots between two and four meters.
- Productive period (reproductive adults) individuals with inflorescences.
- Plants without sexual expression (non-reproductive adults).
2.2. Study Area
2.3. Site distribution in the State of Veracruz 90
2.4. Edaphoclimatic Conditions and Habitat
Georeferencing and Statistical Analysis
3. Results and Discussion
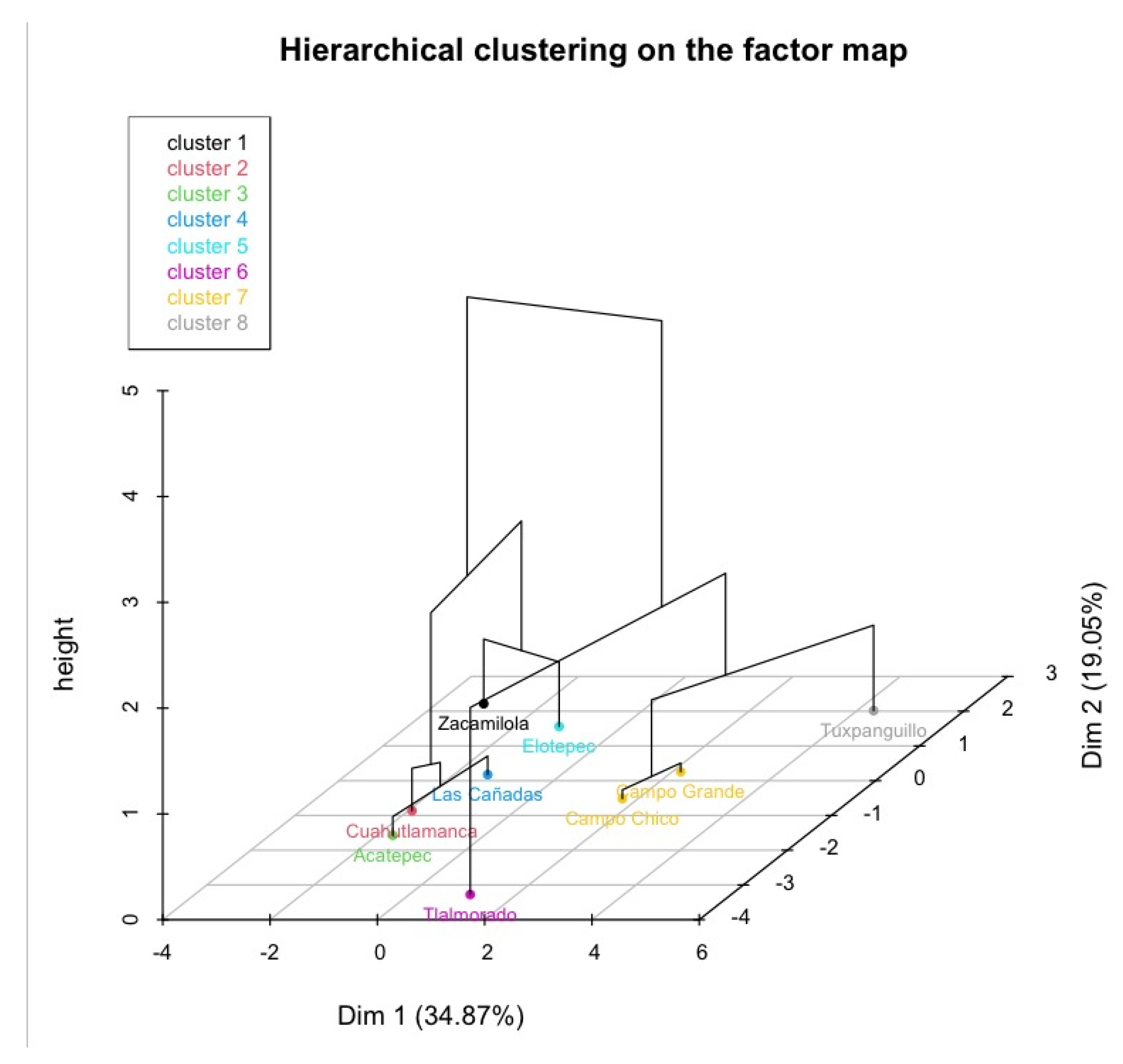
3.1. Distribution of V. tiliifolia
3.2. Edaphoclimatic Conditions and Habitat
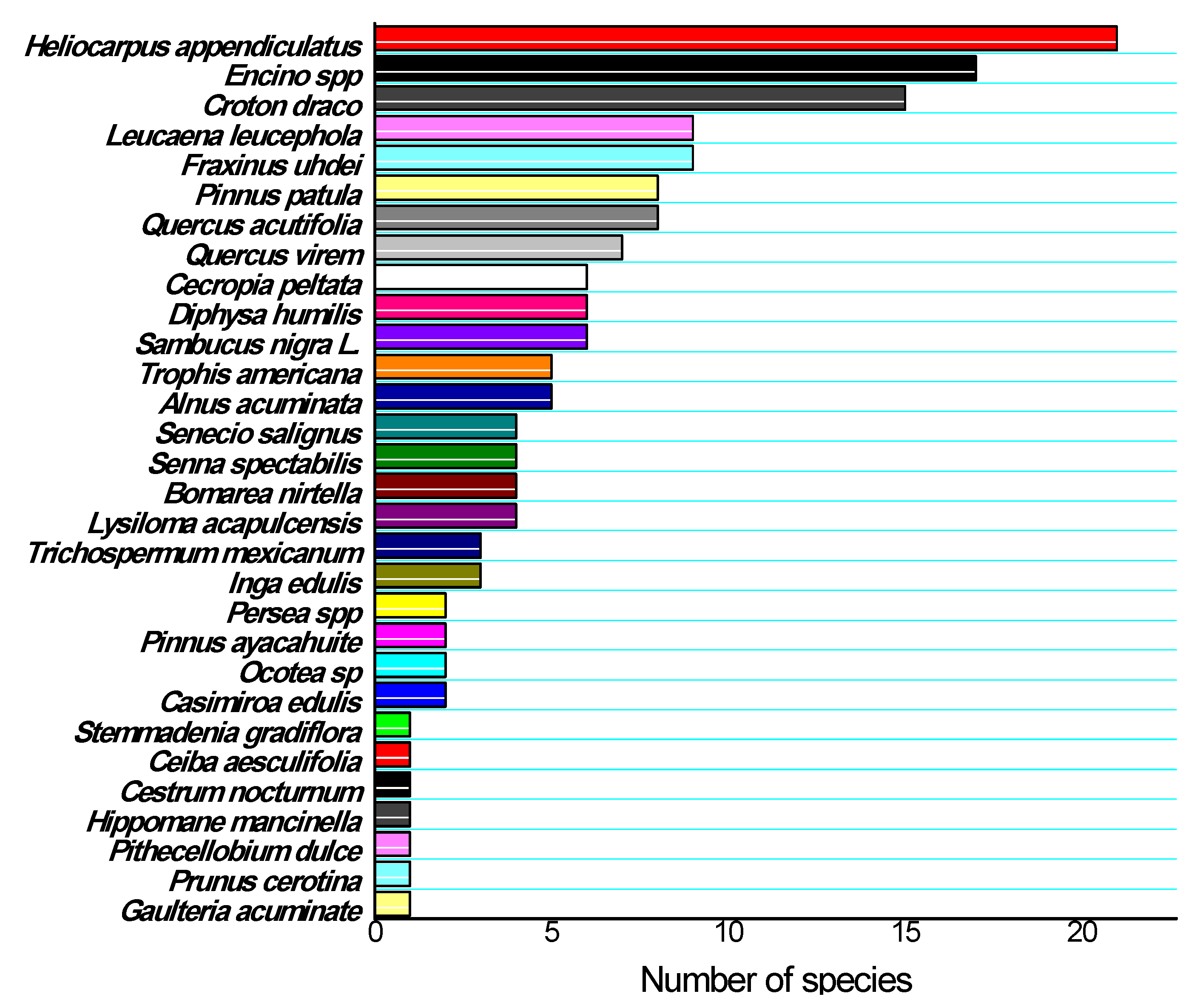
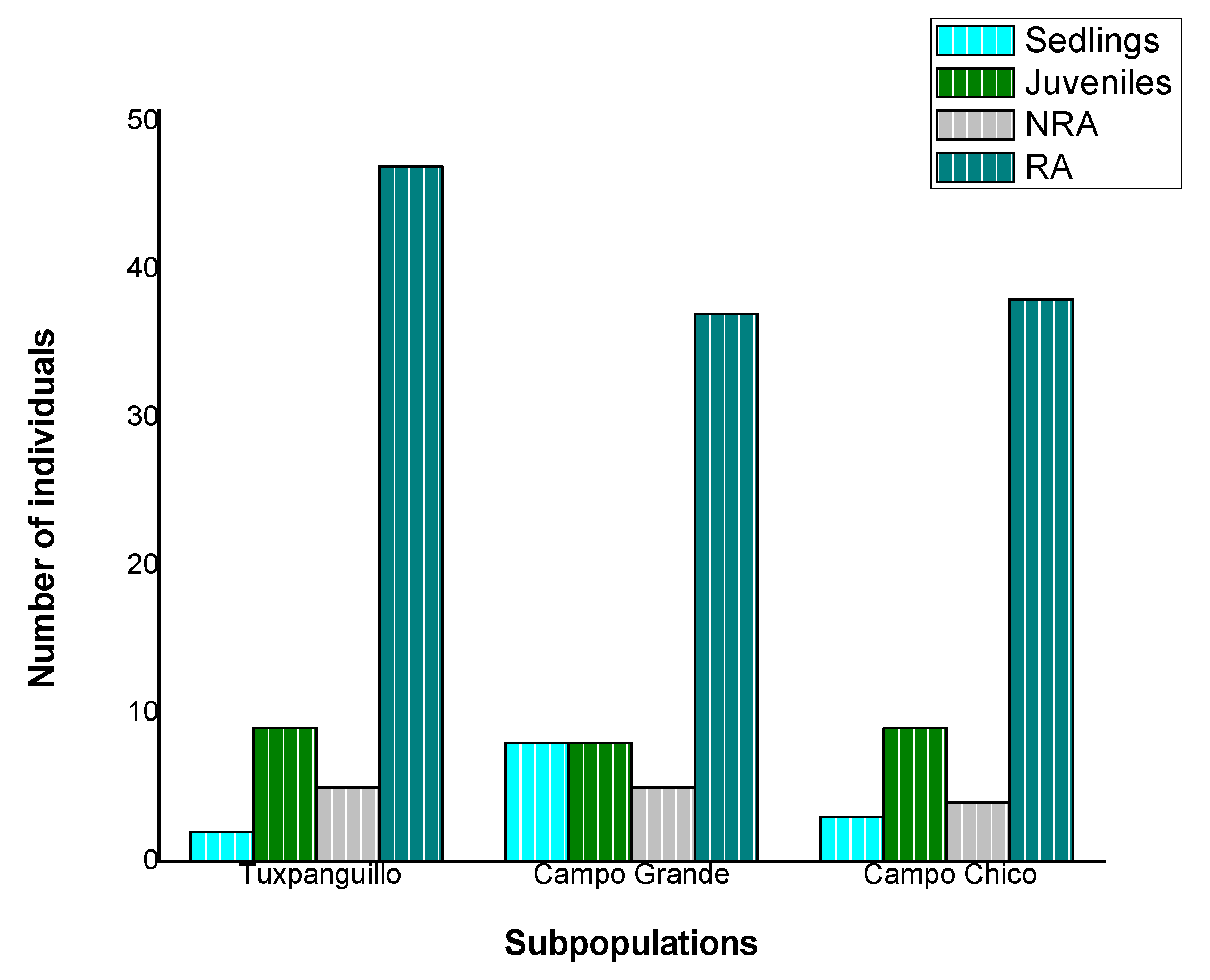
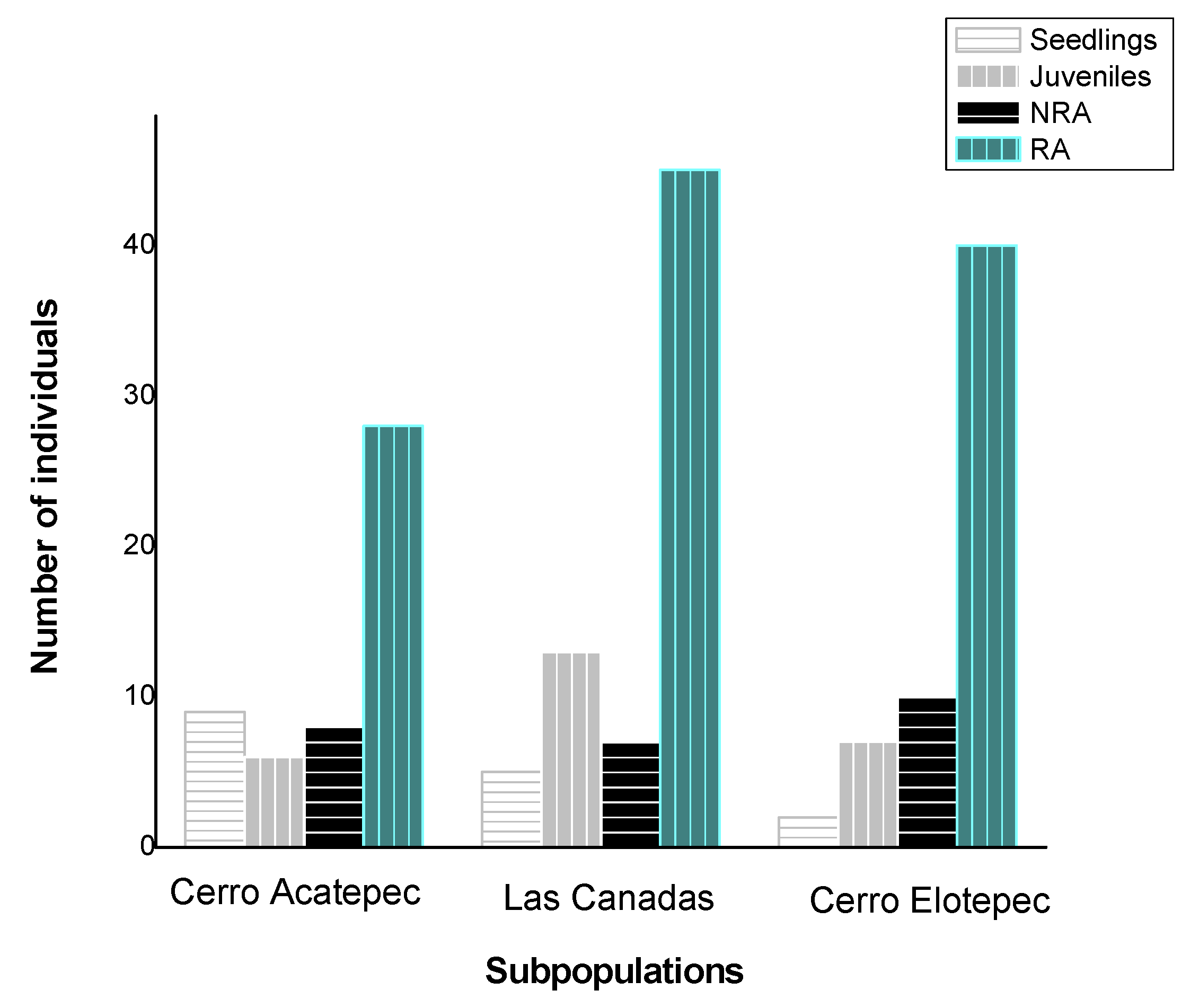
3.3. Sex and Age Structure of the Natural Populations Studied
3.4. Population Density
3.5. Spatial Distribution
3.6. Components of Population Structure and Soil Characteristics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Datos Sobre Alimentación y Agricultura. 2016. Available online: https://www.fao.org/faostat/es/#data/MK (accessed on 11 December 2021).
- Zecca, G.; Jabbott, R.; Sun, W.B.; Spada, A.; Sala, F.; Grassi, F. The timing and the mode of evolution of wild grapes (Vitis). Mol. Phylogenetics Evol. 2012, 66, 736–747. [Google Scholar] [CrossRef] [PubMed]
- SIAP. Anuario Estadístico de la Producción Agrícola en México. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. 2021. Available online: http://nube.siap.gob.mx/cierre_agricola/ (accessed on 5 February 2021).
- Informe Nacional México (FAO). 2006. Estudio de Tendencias y Perspectivas del Sector Forestal en América Latina al Año 2020. Roma. CESOP-IL-72-14-deforestacionEnMexico-310717. Available online: https://www.researchgate.net/publication/308141519_Rojo_Juan_Manuel_Torres_Estudio_de_tendencias_y_perspectivas_del_sector_forestal_en_America_Latina_al_ano_2020_informe_nacional_Mexico (accessed on 12 November 2021).
- Ocete, R.R.; López, M.A.; Gallardo, M.A.; Arnold, C.C.; Pérez, I.M.A.; Rubio, I.M. La Vid Silvestre en el País Vasco y Territorios Limítrofes: Ecología, Distribución y Riesgos para su Conservación; Servicio Central de Publicaciones del Gobierno Vasco: Vitoria-Gazteis, España, 2004; p. 171. [Google Scholar]
- Liang, Z.; Yang, Y.; Cheng, L.; Zhong, G.Y. Polyphenolic composition and content in the ripe berries of wild Vitis species. Food Chem. 2012, 132, 730–738. [Google Scholar] [CrossRef]
- Staudt, G.; Kassemeyer, H.H. Evaluation of downy mildew resistance in various accessions of wild Vitis species. Vitis 1995, 34, 225–228. [Google Scholar]
- Cousins, P. Genetic Control of Phyllotaxy Phase Shift in Juvenile Vines in a Rootstock Hybrid Population. In Proceedings of the 10th International Conference on Grapevine Breeding and Genetics, Geneva, NY, USA, 1–5 August 2014; pp. 275–279. [Google Scholar]
- Galindo-Tovar, M.E.; Davila-Lezama, M.D.R.; Galicia-Sánchez, A.; Olivares-Blanco, E.; Guerra-Ramírez, D.; Aguilar-Rivera, N.; Hernández-Rodríguez, G.; Famiani, F.; Cruz-Castillo, J.G. Artisanal alcoholic beverages made with Vitis tiliifolia grape in Mexico. Rev. Chapingo. Ser. Hortic. 2019, 25, 169–183. [Google Scholar] [CrossRef]
- Tobar-Reyes, J.R.; Franco-Mora, O.; Morales-Rosales, E.J.; Cruz-Castillo, J.G. Phenols of pharmacological interest in wild vines (Vitis spp.) of Mexico. Bol. Latinoam. Caribbean. Med. Aromat. Plants 2011, 10, 167–172. [Google Scholar]
- Jiménez, M.; Juárez, N.; Jiménez, V.M.; Monribot, V.J.L.; Guerrero, A.J.A. Phenolic compounds and antioxidant activity of wild grape (Vitis tiliifolia). Italia. JournalJ Food Science. 2018, 30, 1. [Google Scholar] [CrossRef]
- Juárez, T.N.; Jiménez, F.V.; Guerrero, A.J.A.; Monribot, V.J.L.; Jiménez, F.M. Characterization of oil and flour obtained from wild grape seed (Vitis tiliifolia). Rev. Mex. De Cienc. Agrícolas 2017, 8, 1113–1126. [Google Scholar]
- Medellin-Morales, S.G.; Barrientos-Lozano, L.; Mora-Olivo, A.; Almaguer-Sierra, P.; Mora-Ravelo, S.G. Traditional knowledge and valuation of useful plants in el cielo biosphere reserve, tamaulipas, mexico. Agric. Soc. Y Desarro. 2018, 15, 354–377. [Google Scholar]
- Trujillo, N.J.R.; Fernández, M.J.N.; Analco, J.A.G.; Villanueva, J.L.M.; Fernandez, M.J.N. Caracterización del aceite y harina obtenido de la semilla de uva silvestre (Vitis tiliifolia). Rev. Mex. De Cienc. Agrícolas 2017, 8, 1113–1126. [Google Scholar] [CrossRef] [Green Version]
- Alejandro, H.M.; Cruz Castillo, J.G.; Galindo Tovar, M.E.; Ramirez, D.G.; Famiani, F.; Leyva Ovalle, O.R.; Monribot Villanueva, J.L.; Guerrero Analco, J.A. Phenolic Content and Antioxidant Capacity of Infusions of Vitis tiliifolia (Humb & Bonpl. Ex Schult.) Leaves. J. Agric. Sci. Technol. 2020, 22, 829–836. [Google Scholar]
- Sawda, C.; Moussa, C.; Turner, R.S. Resveratrol for Alzhaimer´s Disease. Ann. N. Y. Acad. Sci. 2017, 1403, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.W.; Kang, H.S. AntiDiabetic Effect of Cotreatment with Quercetin and Resveratrol in Streptozotocin-Induced Diabetic Rats. Biomol. Ther. 2018, 26, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, L. Tratado de Viticultura General, 3rd ed.; Mundi-Prensa: Madrid, Spain, 2002; p. 983. [Google Scholar]
- Gerwing, J.J. Degradación de los bosques por la tala y el fuego en el este de la Amazonía brasileña. Ecol. Gestión For. 2002, 157, 131–141. [Google Scholar]
- Cruz-Castillo, J.G.; Franco-Mora, O.; Famiani, F. Presence and uses of wild grapevines in Central Veracruz, Mexico. J. Int. Sci. Vigne Vin. 2009, 43, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Norma Oficial Mexicana. NMX-AA-132-SCFI-2006. Muestreo de Suelos para la Identificación y la Cuantificación de Metales y Metaloides, y Manejo de la Muestra. Available online: https://www.cmic.org.mx/comisiones/Sectoriales/medioambiente/Varios/Leyes_y_Normas_SEMARNAT/NMX/Contaminaci%C3%B3n%20del%20Suelo/2.2006.pdf (accessed on 15 October 2021).
- Vázquez, T.M.; Campos., J.J.; Juarez, F.M.A. Tropical Trees of Veracruz; Universidad Veracruzana Editorial Direction: Xalapa, Mexico, 2017; p. 517. [Google Scholar]
- National Commission for the Use of Biodiversity, Conabio (2020) Biodiversity. Mexico. Available online: https://www.biodiversidad.gob.mx/biodiversidad/ (accessed on 13 January 2022).
- IUSS, Grupo de Trabajo WRB. 2007. Base Referencial Mundial del Recurso Suelo. Primera Actualización 2007. Informes Sobre Recursos Mundiales de Suelos. FAO, Roma. 2007. 103 p. Available online: https://www.fao.org/3/a0510s/a0510s00.pdf (accessed on 22 June 2021).
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Englewood Cliffs, NJ, USA, 1988; p. 718. [Google Scholar]
- Servicio de Inspección and Certificación de Semillas. (SNICS). 2017. Red de la Vid (Vitis tiliifolia H). Available online: https://www.gob.mx/snics/acciones-y-programas/vid-vitis-tiliifolia-h (accessed on 22 June 2021).
- Rzedowski, J.; Calderón, G.R. Flora of the Bajío and Adjacent Regions. Fascicle 131. Vitacea; Institute of Ecology: Pátzcuaro, México, 2005; p. 31. [Google Scholar]
- Villaseñor, J.; Ortiz, E. The Asteraceae family in the flora of the Bajío and adjacent regions. Mex. Bot. Act 2013, 100, 263–295. [Google Scholar]
- Geertje, M.F.; van der Heijden, L.; Phillips, O. What controls liana success in Neotropical forests? Glob. Ecol. Biogeogr. 2008, 17, 372–383. [Google Scholar]
- Schnitzer, S.A.; Bongers, F. The ecology of lianas and their role in forests. Trends Ecol. Evol. 2002, 17, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Salicrup, D.R.; Barker, M.G. Effect of liana cutting on water potential and growth of adult Senna multijuga (Caesalpinioideae) trees in a Bolivian tropical forest. Oecologia 2000, 124, 469–475. [Google Scholar] [CrossRef]
- Castillo-Campos, G.; Avendaño, S.; Medina, M.E. Flora y vegetación. In La Biodiversidad en Veracruz: Estudio de Estado; Cruz-Angón, A., Ed.; CONABIO. Gobierno del Estado de Veracruz, Universidad Veracruzana, Instituto de Ecología, A.C.: Xalapa, México, 2011; pp. 163–179. [Google Scholar]
- Wright, S.J.; Jaramillo, M.A.; Pavón, J.; Condit, R.; Hubbell, S.P.; Foster, R.B. Reproductive size thresholds in tropical trees: Variation among individuals, species and forests. J. Trop. Ecol. 2005, 21, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Kainer, K.A.; Wadt, L.H.O.; Gomes-Silva, D.A.P.; Capanu, M. Liana loads and their association with Bertholletia excelsa fruit and nut production, diameter growth and crown attributes. J. Trop. Ecol. 2006, 22, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Schnitzer, S.A.; Carson, W.P. Lianas suppress tree regeneration and diversity in treefall gaps. Ecol. Lett 2010, 13, 849–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dillenburg, L.R.; Whigham, D.F.; Teramura, A.H.; Forseth, I.N. Effects of below and above ground competition from the vines Lonicera japonica and Parthenocissus quinquifolia on the growth of the host Liquidambar styraciflua. Oecologia 1993, 93, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Manríquez, G.; Rendón-Sandoval, F.J.; Cornejo-Tenorio, G.; Carrillo-Reyes, P. Lianas of Mexico. Bot. Sci. 2015, 93, 365–417. [Google Scholar] [CrossRef] [Green Version]
- Matallana, G.; Wendt, T.; Araujo, D.S.; Scarano, F.R. High abundance of dioecious plants in a tropical coastal vegetation. Am. J. Bot. 2005, 92, 1513–1519. [Google Scholar] [CrossRef] [PubMed]
- Primack, R.B. A Primer of Conservation Biology; Sinauer Associates: Sunderland, MA, USA, 1995. [Google Scholar]
- Poorter, L.; Bongers, F.; Van Rompaey, R.S.A.R.; De Klerk, M. Regeneration of canopy tree species at five sites in West African moist forest. For. Ecol. Manag. 1996, 84, 61–69. [Google Scholar] [CrossRef]
- Sosa, V.J.; Vovides, A.P.; Castillo-Campos, G. Monitoring endemic plant extinction in Veracruz, Mexico. Biodiver. Conserv. 1998, 7, 1521–1527. [Google Scholar] [CrossRef]
- George, A.K.; Walker, K.F.; Lewis, M.M. Population status of eucalypt trees on the river Murray floodplain, South Australia. Riv. Res. App. 2005, 21, 271–282. [Google Scholar] [CrossRef]
- Leak, W.B. The Jshaped probability distribution. For. Sci. 1965, 11, 405–409. [Google Scholar] [CrossRef]
- Bierzychudek, P. Introduction to plant population ecology. In Trends in Ecology and Evolution, 2nd ed.; Elsevier: Victory, Austalia, 1987. [Google Scholar] [CrossRef]
- Lamprecht, H. Silvicultura en Los Trópicos. Los Ecosistemas Forestales en los Bosques Tropicales y sus Especies Arbóreas. Posibilidades para un Aprovechamiento Sostenido. Eschborn: GTZ. Deutsche Gesellschaft für Technische Zusammenarbeit. 1990; pp. 288–292. 1990, p. 288. Available online: https://bibliotecadigital.infor.cl/handle/20.500.12220/1232 (accessed on 13 January 2022).
- Howard, A.F.; Valerio, J. A diameter class growth model for assessing the sustainability of silvicultural prescriptions in natural tropical forests. Commonw. For. Rev. 1992, 71, 171–177. [Google Scholar]
- Meléndez, E.; Puras, P.; García, J.L.; Cantos, M.; Gómez-Rodríguez, J.A.; Íñiguez, M.; Rodríguez Álvaro Valle, J.M.; Arnold, C.; Ocete, C.A.; Ocete, R. Evolución de las vides silvestres y asilvestradas del bosque de galería del río Ega (País Vasco y Navarra, España) desde 1995 hasta 2015. OENO One 2016, 50, 65–75. [Google Scholar] [CrossRef]
- Silvertown, J.W.; Lovett-Doust, J. Introduction to Plant Population Ecology; Blackwell Scientific Publications: Oxford, UK, 1993; p. 210. [Google Scholar]
- Smith, D.M.; Larson, B.C.; Kelty, M.J.; Ashton, P.M.S. The Practice of Silviculture: Applied Forest Ecology; John Wiley & Sons: New York, NY, USA, 1997. [Google Scholar]
- Wright, S.J.; Muller-Landau, H.C.; Condit, R.; Hubbell, S.P. Gap-dependent recruitment, realized vital rates, and size distributions of tropical trees. Ecology 2003, 84, 3174–3185. [Google Scholar] [CrossRef]
- Souza, A.F. Ecological interpretation of multiple population size structures in trees: The case of Araucaria angustifolia in South America. Austral Ecol. 2007, 32, 524–533. [Google Scholar] [CrossRef]
- Ballina-Gómez, H.S.; Iriarte-Vivar, S.; Orellana, R.; Santiago, L.S. Respuestas de crecimiento compensatorio a la defoliación y disponibilidad de luz en dos especies de plantas leñosas nativas de México. Rev. De Ecol. Trop. 2010, 26, 163–171. [Google Scholar] [CrossRef]
- Mascaro, J.; Schnitzer, S.A.; Carson, W.P. Liana diversity, abundance, and mortality in a wet tropical forest. Forest. Ecol. Manag. 2004, 190, 3–14. [Google Scholar] [CrossRef] [Green Version]
- DeWalt, S.J.; Schntizer, S.A.; Denslow, J.S. Density and diversity of lianas along a chronosequense in a central Panamanian lowland forest. J. Trop. Ecol. 2000, 16, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Schnitzer, S.A.; DeWalt, S.J.; Chave, J. Censusing and measuring lianas: A quantitative comparison of the common methods. Biotropica 2006, 38, 581–591. [Google Scholar] [CrossRef]
- Strasburger, E.; Noll, F.; Schenck, H.; Schimper, A.; Franz, W. Treaty of Botany, 35th ed.; Sitt, P., Ed.; Omega: Barcelona, Spain, 2004. [Google Scholar]
- Norma Oficial Méxicana. NOM-059-SEMARNAT 2010. Available online: http://dof.gob.mx/nota_detalle_popup.php?codigo=5173091 (accessed on 12 September 2021).
- Biagini, B.; Imazio, S.; Scienza, A.; Failla, O.Y.; De Lorenzis, G. Renovación de las poblaciones de vid silvestre (Vitis vinifera L. subsp. sylvestris (Gmelin) Hegi) a través de la vía sexual: Algunos estudios de casos italianos. Flora 2016, 219, 85–93. [Google Scholar] [CrossRef]
- Bonguer, F.; Popma, J.; Meave Del Castillo, J.; Carabias, J. Structure and floristic composition of the lowland rain forest of The Tuxtlas, Mexico. Vegetatio 1988, 74, 55–80. [Google Scholar] [CrossRef]
- Putz, F.E. Climber Ecology. Eco. Info 23 2004. Available online: www.ecologia.info/trepadoras.htm (accessed on 13 July 2021).
- Deslow, J.S.; Guzman, G.S. Variation in stand structure, ligh and seedling abundance, a cross a tropical moist forest cronosequence, Panama. J. Veg. Sci. 2000, 11, 201–212. [Google Scholar] [CrossRef]
- Aguirre, O.A.; Hui, G.; Gadow, K.; Jiménez, J. An analysis of spatial forest structure using neighbourhood-based variables. For. Ecol. Manag. 2003, 183, 137–145. [Google Scholar] [CrossRef]
- Pommerening, A. Approaches to quantifying forest structures. J. For. 2002, 75, 305–324. [Google Scholar] [CrossRef]
- Schneider, A.; Mainarde, E.; Raimondi, S. Ampelography Universale Storica Illustrata. L´Artistica Editrice; Centro Studi Piemontesi: Turin, Italy, 2012. [Google Scholar]
- Martínez-Ramos, M.; Álvarez-Buylla, E.R. Ecology of plant populations in a humid forest of Mexico. Bull. Bot. Soc. Mex. 1995, 56, 121–153. [Google Scholar] [CrossRef] [Green Version]
- Arnáez, J.; Ortigosa, L.; Ruiz-Flaño, P.; Lasanta, T. Sediment production in vineyards through rainfall simulation: Comparison of results with different empirical models. In Geomorphology and Territory; Pérez Alberti, A., López Bedoya, J., Eds.; Publications and Scientific Exchange Service of the University of Santiago de Compostela: Santiago de Compostela, España, 2006; pp. 165–175. [Google Scholar]
- Summer, M.E.; Fey, M.V.; Noblen, D. Nutrient Stratusand and Toxity Problems and Acid Soils; En, B., Ullrich, M.E., Summer, M.E., Eds.; Springuer-Verlag: Berlin, Germany, 1991; pp. 149–182. [Google Scholar]
- Rog-Young, K.; Jeong-Ki, Y.; Tae-Seung, K.; Jae, Y.; Gary, O.; Kwon-Rae, K. Bioavailability of heavy metals in soils: Definitions and practical implementation—a critical review. Environ. Geochem. Health 2015, 37, 1041–1061. [Google Scholar] [CrossRef]
- Porro, D.; Steffanini, M.; Failla, O.; Stringari, G. Optimal leaf sampling time in diagnosis of grapevine nutritional status. Acta Hort. 1995, 383, 135–142. [Google Scholar] [CrossRef]
- Bonomelli, C.; Ruiz, R. Effects of foliar and soil calcium application on yield and quality of table grape cv. “Thompson Seedless”. J. Plant Nutr. 2010, 33, 299–314. [Google Scholar] [CrossRef]
- Downton, W.J.S. Crecimiento y composición mineral de la vid sultana según la influencia de la salinidad y el patrón. Rev. Aust. De Investig. Agrícola 1985, 36, 425–434. [Google Scholar]
- Walker, R.R.; Blackmore, D.H.; Clingeleffer, P.R.; Iacono, F. Efecto de la salinidad y el portainjertos Ramsey sobre las concentraciones de iones y la asimilación de dióxido de carbono en hojas de vid cultivadas en el campo con riego por goteo (Vitis vinifera L. cv. Sultana). Rev. Aust. De Investig. Sobre La Uva Y El Vino 1997, 3, 66–74. [Google Scholar]
- Fisarakis, I.; Chartzoulakis, K.; Stavrakas, D. Respuesta de las vides Sultana (V. vinifera L.) en seis portainjertos a la exposición y recuperación de la salinidad del NaCl. Gestión Del Agua Agrícola 2001, 51, 13–27. [Google Scholar]
- Massai, R.; Remorini, D.; Tattini, M. Intercambio de gases, relaciones hídricas y ajuste osmótico en dos combinaciones de vástagos/portainjertos de Prunus bajo varias concentraciones de salinidad. Planta Y Suelo. 2004, 259, 153–162. [Google Scholar] [CrossRef]
- Vila, H.F.; Di Filippo, M.L.; Venier, M.; Hugalde, I.P.; Filippini, M.F. A dynamic model for sodium intoxication unravels salt tolerance in grapevine (Vitis vinifera L.) rootstocks. Rev. Fac. Cienc. Agrarias. Univ. Nac. Cuyo 2020, 52, 88–101. [Google Scholar]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.; Thuleau, P.; Mazars, C. Calcium sensors as key hub in plant resses to biotic and abiotic stresses. Front. Plant Sci. 2016, 16, 327. [Google Scholar]
- Maya-Meraz, I.; Pérez-Leal, R. ¿Qué relación existe entre la calidad aromática del vino y la bioquímica de la vid? Tecnociencia Chihuah. 2019, 12, 134–142. [Google Scholar]
- Mata.Alejandro, H.; Galindo-Tovar, M.E.; Leyva-Ovalle, O.; Famiani, F.; Cruz-Castillo, J.G. Environmental Conditions, and Phenolic Compounds Potential in the Leaves of Vitis Tiliifolia; Veracruz, México. 2021. Available online: https://www.researchsquare.com/article/rs-183717/v1 (accessed on 22 June 2021).
- Marshner, H. Mineral Nutrition of Highger Plants, 2nd ed.; Academic Press: London, UK, 1996. [Google Scholar]
- Miyamoto, K.; Suzuki, E.; Kohyama, T.; Seino, T.; Mirmanto, E.; Simbolon, H. Habitat differentiation among tree species with small-scale variation of humus depth and topography in a tropical heath forest of Central Kalimantan, Indonesia. J. Trop. Ecol. 2003, 19, 43–54. [Google Scholar] [CrossRef]
- Philips, O.; Nuñez-Vargas, L.; Lorenzo-Monteagudo, A.P.; Peña-Cruz, A.M.; Chuspe-Zans, E.W.; Galiano-Sanchez, M.; Yli-Halla, R.S. Habitat association among Amazonian tree species: Alandscape-scale approach. J. Ecol. 2003, 91, 757–775. [Google Scholar] [CrossRef] [Green Version]
- Tuomisto, H.; Ruokolainen, K.; Yli–halla, M. Dispersal, environment, and floristic variation of western amazonian forests. Science 2003, 299, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.S.; MCkenna, J.J.; Ashton, P.M.D.; Gregoire, T.G. Habitat characterization underestimates the role of edaphic factors controlling the distribution of Entandrophragma. Ecology 2004, 85, 2171–2183. [Google Scholar] [CrossRef]
- Paoli, G.D.; Curran, L.M.; Zak, D.R. Soil nutrients and beta diversity in the Bornean Dipterocarpaceae: Evidence for niche partitioning by tropical rain forest trees. J. Ecol. 2006, 94, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Balfour, D.A.; Bond, W.J. Factors limiting climber distribution and abundance in a southern African forest. J. Ecol. 1993, 81, 93–99. [Google Scholar] [CrossRef]
- DeWalt, S.J.; Chave, J. Structure and biomass of four lowland neotropical forests. Biotropica 2004, 36, 7–19. [Google Scholar] [CrossRef]
- Macía, M.; Ruokolainen, K.; Tuomisto, H.; Quisbert, J.; Cala, V. Congruence between floristic patterns of trees and lianas in a southwest Amazonian rain forest. Ecography 2007, 30, 561–577. [Google Scholar] [CrossRef]
- Graham, E.A.; Mulkey, S.S.; Kitajima, K.; Phillips, N.G.; Wright, S.J. Cloud cover limits net CO2 uptake and growth of a rainforest tree during tropical rainy seasons. Proc. Natl. Acad. Sci. USA 2003, 100, 572–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.Q.; Pooter, L.; Cao, K.F.; Bonguers, F. Seedling Growth Strategies in Bauhinia Species: Comparing Lianas and Trees. Ann. Bot. 2007, 100, 831–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Aust. J. Plant Physiol. 2000, 27, 595–607. [Google Scholar] [CrossRef] [Green Version]
- Poorter, L. Resource capture and use by tropical forest tree seedlings and their consequences for competition. In Biotic Interactions in the Tropics: Their Role in the Maintenance of Species Diversity; Burslem, D.F.R.P., Pinard, M.A., Hartley, S.E., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 35–64. [Google Scholar] [CrossRef]
- Arrey-Salas, O.; CarisMaldonado, J.C.; Hernández-Rojas, B.; Gonzalez, E. Comprehensive Genome-Wide Exploration of C2H2 Zinc Finger Family in Grapevine (Vitis vinifera L.): Insights into the Roles in the Pollen Development Regulation. Genes 2021, 12, 302. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Municipality | Coordinates | Altitude (m.a.s.l.) | Localities |
|---|---|---|---|
| Huatusco | 19°04′ and 19°13′ N 96°41′ and 97° 04′ W | 400–2000 | Cerro Elotepec |
| Cerro Acatepec | |||
| Las Cañadas | |||
| Ixtaczoquitlán | 18°45′ and 18°57′ N 96°58′ and 97°06′ W | 700–1700 | Tuxpanguillo |
| Campo Chico | |||
| Campo Grande | |||
| Atlahuilco | 18°38′ and 18°44′ N 97°03′and 97°11′ W | 1760–2700 | Cuahutlamanca |
| Tlalmorado | |||
| Zacamilola |
| Herbarium | Xal-INECOL, Xalapa Rzedowski Faculty of Biological and Agricultural Sciences, UV Peñuela, Cordoba. CORU Herbarium of the Faculty and Agricultural Sciences of the University Veracruzana, campus Xalapa, Veracruz |
| Databases | Tropics (Missouri Botanical Garden) Botanical Society of México Virtual Herbarium CONABIO. National Commission for the Knowledge and Use of Biodiversity |
| Ixtaczoquitlán | Huatusco | Atlahuilco | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Year | °T | PP | RH | °T | PP | RH | °T | PP | RH |
| 2000 | 18.3 | 209.4 | 81 | 23.10 | 170.23 | 18.3 | 209.4 | 81 | |
| 2001 | 18.56 | 190.5 | 82 | 22.86 | 161 | 18.56 | 190.5 | 82 | |
| 2002 | 18.72 | 142.9 | 82 | 23.58 | 160.34 | 18.72 | 142.9 | 82 | |
| 2003 | 19.03 | 237.22 | 83 | 22.76 | 154.49 | 19.03 | 237.22 | 83 | |
| 2004 | 18.66 | 166.55 | 79 | 22.76 | 154.49 | 18.66 | 166.55 | 79 | |
| 2005 | 19.03 | 169.50 | 78 | 23.02 | 165 | 19.03 | 169.50 | 78 | |
| 2006 | 19.02 | 188.61 | 74 | 22.32 | 166.42 | 19.02 | 188.61 | 74 | |
| 2007 | 18.97 | 186.06 | 81 | 22.26 | 127.2 | 18.97 | 186.06 | 81 | |
| 2008 | 18.97 | 194.12 | 79 | 22.50 | 156.59 | 18.97 | 194.12 | 79 | |
| 2009 | 19.21 | 171.91 | 75 | 22.70 | 162.63 | 19.21 | 171.91 | 75 | |
| 2010 | 18.73 | 211.58 | 80 | 22.33 | 172 | 18.73 | 211.58 | 80 | |
| 2011 | 19.40 | 188.45 | 77 | 22.79 | 149.38 | 19.40 | 188.45 | 77 | |
| 2012 | 19.07 | 231.03 | 80 | 22.56 | 185.5 | 19.07 | 231.03 | 80 | |
| 2013 | 19.33 | 210.33 | 80 | 22.35 | 197.6 | 19.33 | 210.33 | 80 | |
| 2014 | 19.07 | 204.37 | 80 | 22.70 | 200.42 | 19.07 | 204.37 | 80 | |
| 2015 | 19.42 | 197.22 | 81 | 22.73 | 166.24 | 19.42 | 197.22 | 81 | |
| 2016 | 19.71 | 225.92 | 79 | 23.33 | 171.25 | 19.71 | 225.92 | 79 | |
| 2017 | 19.67 | 187.23 | 79 | 23.42 | 158.21 | 19.67 | 187.23 | 79 | |
| Category | Huatusco Index | Ixtaczoquitlán Index | Atlahuilco Index | Distribution Pattern | |||
|---|---|---|---|---|---|---|---|
| Seedlings | H1 | 0.26 | I1 | 0.67 | A1 | Np | Aggregation |
| H2 | 0.40 | I2 | 0.38 | A2 | 0.24 | Aggregation | |
| H3 | 0.45 | I3 | 0.30 | A3 | 0.37 | Aggregation | |
| Juvenile | H1 | 0.39 | I1 | 0.60 | A1 | 0.43 | Aggregation |
| H2 | 0.44 | I2 | 0.35 | A2 | 0.17 | Aggregation | |
| H3 | 0.33 | I3 | 0.37 | A3 | 0.25 | Aggregation | |
| Reproductive adults | H1 | 0.97 | I1 | 1.23 | A1 | 1.19 | Random |
| H2 | 1.19 | I2 | 0.76* | A2 | 0.5* | Random | |
| H3 | 0.93 | I3 | 1.07 | A3 | 0.79* | Random | |
| Municipality | Site | pH | OM (%) | CEC Cmol.Kg−1 | CaCO3 (%) | N (%) | P mg/kg−1 | K mg/kg−1 | Ca mg/kg−1 |
|---|---|---|---|---|---|---|---|---|---|
| H | Acatepec | 5.9 | 8.5 | 15.35 | 0.13 | 0.256 | 1.34 | 159.9 | 1936.8 |
| H | Elotepec | 6 | 7 | 14.28 | 0.77 | 0.244 | 6.34 | 127 | 1327 |
| H | Las Cañadas | 5.3 | 8.1 | 17.81 | 0.38 | 0.35 | 3.12 | 164.9 | 2875 |
| I | Campo Chico | 5.8 | 5 | 19 | 0.28 | 0.174 | 3.02 | 116.9 | 2549.23 |
| I | Campo Grande | 6.1 | 6.1 | 16.7 | 0.58 | 0.195 | 2.45 | 123.81 | 1980 |
| I | Tuxpanguillo | 6.21 | 7.5 | 24.64 | 0.75 | 0.241 | 3.19 | 106.08 | 3582.6 |
| A | Cuahutlamanca | 5.7 | 9 | 17.02 | 0.59 | 0.25 | 5.2 | 198 | 1500 |
| A | Tlalmorado | 6.3 | 7.2 | 15.8 | 0.41 | 0.169 | 6.7 | 159.04 | 3581 |
| A | Zacamilola | 5.2 | 6.7 | 13.26 | 0.81 | 0.27 | 4.9 | 133.76 | 2369 |
| Municipality | Site | Mg mg/kg−1 | Na mg/kg−1 | Fe mg/kg−1 | Cu mg/kg−1 | Zn mg/kg−1 | Mn mg/kg−1 | B mg/kg−1 | |
| H | Acatepec | 174.34 | 32.43 | 28.06 | 0.39 | 1.66 | 28.12 | 0.99 | |
| H | Elotepec | 254.7 | 82.4 | 54.3 | 1.64 | 2.54 | 15.2 | 0.73 | |
| H | Las Cañadas | 101.23 | 42.5 | 68.12 | 1.04 | 4.2 | 9.58 | 0.49 | |
| I | Campo Chico | 289.6 | 23.12 | 83.6 | 0.49 | 4.2 | 6.5 | 0.57 | |
| I | Campo Grande | 341.7 | 28.9 | 101.3 | 0.97 | 6.3 | 18.3 | 0.65 | |
| I | Tuxpanguillo | 498.86 | 13.11 | 113.86 | 1.58 | 5.53 | 41.74 | 0.64 | |
| A | Cuahutlamanca | 102.5 | 46.03 | 67 | 1.01 | 2.8 | 17.3 | 0.92 | |
| A | Tlalmorado | 145.8 | 22.6 | 62 | 0.53 | 6.7 | 12.84 | 1.34 | |
| A | Zacamilola | 99.4 | 74.2 | 21.6 | 0.22 | 1.95 | 39.3 | 0.32 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mata-Alejandro, H.; Betanzo-Torres, E.A.; Navarrete-Rodriguez, G.; Castañeda-Chavez, M.D.R. Population Structure of Three Provenances of Vitis tiliifolia (Humb. & Bonpl. ex Schult.) in the Central Zone of the State of Veracruz, México. Agriculture 2022, 12, 659. https://doi.org/10.3390/agriculture12050659
Mata-Alejandro H, Betanzo-Torres EA, Navarrete-Rodriguez G, Castañeda-Chavez MDR. Population Structure of Three Provenances of Vitis tiliifolia (Humb. & Bonpl. ex Schult.) in the Central Zone of the State of Veracruz, México. Agriculture. 2022; 12(5):659. https://doi.org/10.3390/agriculture12050659
Chicago/Turabian StyleMata-Alejandro, Humberto, Erick Arturo Betanzo-Torres, Gabycarmen Navarrete-Rodriguez, and Maria Del Refugio Castañeda-Chavez. 2022. "Population Structure of Three Provenances of Vitis tiliifolia (Humb. & Bonpl. ex Schult.) in the Central Zone of the State of Veracruz, México" Agriculture 12, no. 5: 659. https://doi.org/10.3390/agriculture12050659