
A Review of the Botany, Volatile Composition, Biochemical and Molecular Aspects, and Traditional Uses of Laurus nobilis


Abstract
:1. Introduction
2. Botany and Distribution
3. Origin, Domestication, and Spread
4. Cultivation and Conservation
4.1. Growth Conditions
4.2. Propagation
4.3. Pests and Diseases
4.4. Harvesting
5. Plant Genetic Resources
6. Characterization and Evaluation
6.1. Characterization for Essential Features and Classification
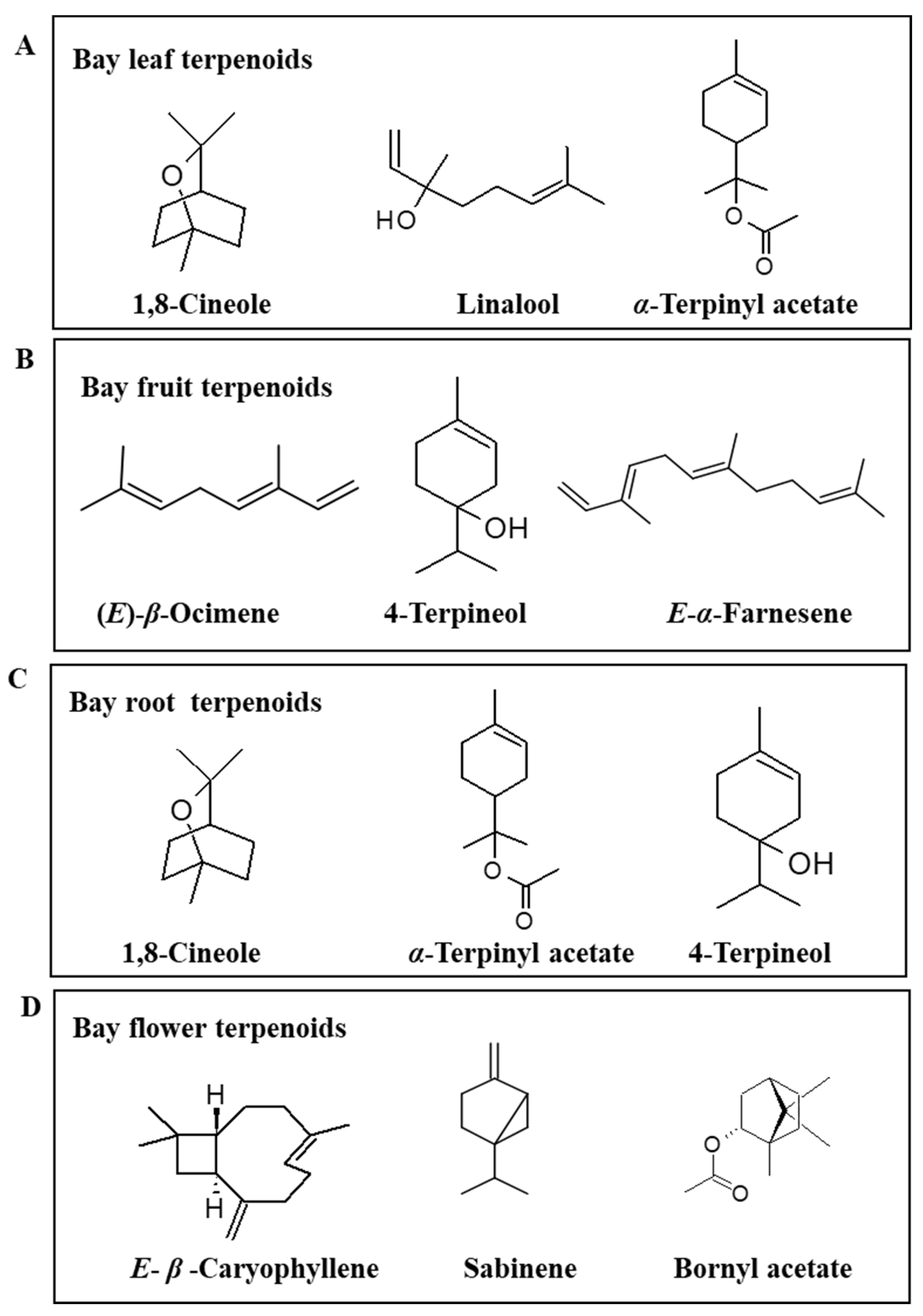
6.2. Characterization of Laurus nobilis for its Essential Oil and Volatile Composition
6.2.1. Leaf
6.2.2. Fruit
6.2.3. Root
6.2.4. Flower
7. Factors Affecting the Volatile Composition in L. nobilis
7.1. Extraction Methods
7.2. Drying Methods
7.3. Season
8. Molecular Characterization
8.1. Molecular Markers
8.2. Identification and Characterization of Novel Genes
8.3. Sex Determination
9. Biological Activity of L. nobilis Essential Oil
9.1. Antioxidant Activity
9.2. Antimicrobial Activity
9.3. Effect on Arthropodos
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dobroslavić, E.; Repajić, M.; Dragović-Uzelac, V.; Elez Garofulić, I. Isolation of Laurus nobilis leaf polyphenols: A Review on current techniques and future perspectives. Foods 2022, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- Tucker, A.O.; DeBaggio, T. The Encyclopedia of Herbs: A Comprehensive Reference to Herbs of Flavor and Fragrance; Timber Press: Portland, OR, USA, 2009. [Google Scholar]
- Sangun, M.K.; Aydin, E.; Timur, M.; Karadeniz, H.; Caliskan, M.; Ozkan, A. Comparison of chemical composition of the essential oil of Laurus nobilis L. leaves and fruits from different regions of Hatay, Turkey. J. Environ. Biol. 2007, 28, 731–733. [Google Scholar] [PubMed]
- Anzano, A.; de Falco, B.; Grauso, L.; Motti, R.; Lanzotti, V. Laurel, Laurus nobilis L.: A review of its botany, traditional uses, phytochemistry and pharmacology. Phytochem. Rev. 2022, 1–51. [Google Scholar] [CrossRef]
- Ehrendorfer, F.; Krendl, F.; Habeler, E.; Sauer, W. Chromosome numbers and evolution in primitive angiosperms. Taxon 1968, 17, 337–353. [Google Scholar] [CrossRef]
- Sharma, A.; Singh, J.; Kumar, S. Bay leaves. In Handbook of Herbs and Spices (Second Edition), Volume 1; Elsevier: Amsterdam, The Netherlands, 2012; pp. 73–85. [Google Scholar]
- Schultze-Motel, J.; Fritsch, R.; Hammer, K.; Hanelt, P.; Kruse, J.; Maass, H.I.; Ohle, H.; Pistrick, K. Taxonomy and evolution of cultivated plants: Literature review 1984/1985. Die Kulturpflanze 1986, 34, 335–347. [Google Scholar] [CrossRef]
- Yılmaz, B.; Deniz, İ. The effects of cultivation area and altitude variation on the composition of essential oil of Laurus nobilis L. grown in eastern, Western and Central Karadeniz Region. Int. J. Second. Metab. 2017, 4, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Demir, V.; Gunhan, T.; Yagcioglu, A.; Degirmencioglu, A. Mathematical modelling and the determination of some quality parameters of air-dried bay leaves. Biosyst. Eng. 2004, 88, 325–335. [Google Scholar] [CrossRef]
- Sekeroglu, N.; Koca, U. Current status of medicinal and aromatic plants production and trade in Turkey. Biomed 2010, 5, 65–73. [Google Scholar]
- Flamini, G.; Bader, A.; Cioni, P.L.; Katbeh-Bader, A.; Morelli, I. Composition of the essential oil of leaves, galls, and ripe and unripe fruits of Jordanian Pistacia palaestina Boiss. J. Agric. Food Chem. 2004, 52, 572–576. [Google Scholar] [CrossRef]
- Kilic, A.; Hafizoglu, H.; Kollmannsberger, H.; Nitz, S. Volatile constituents and key odorants in leaves, buds, flowers, and fruits of Laurus nobilis L. J. Agric. Food Chem. 2004, 52, 1601–1606. [Google Scholar] [CrossRef]
- Sırıken, B.; Yavuz, C.; Güler, A. Antibacterial activity of Laurus nobilis: A review of literature. Med. Sci. Discov. 2018, 5, 374–379. [Google Scholar] [CrossRef]
- Özcan, M.; Chalchat, J.-C. Effect of different locations on the chemical composition of essential oils of laurel (Laurus nobilis L.) leaves growing wild in Turkey. J. Med. Food 2005, 8, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Ordoudi, S.A.; Papapostolou, M.; Nenadis, N.; Mantzouridou, F.T.; Tsimidou, M.Z. Bay Laurel (Laurus nobilis L.) essential oil as a food preservative source: Chemistry, quality control, activity assessment, and applications to olive industry products. Foods 2022, 11, 752. [Google Scholar] [CrossRef] [PubMed]
- Charles, D.J. Bay. In Antioxidant properties of spices, herbs and other sources; Springer: Berlin/Heidelberg, Germany, 2012; pp. 181–187. [Google Scholar]
- Al-Kalaldeh, J.Z.; Abu-Dahab, R.; Afifi, F.U. Volatile oil composition and antiproliferative activity of Laurus nobilis, Origanum syriacum, Origanum vulgare, and Salvia triloba against human breast adenocarcinoma cells. Nutr. Res. 2010, 30, 271–278. [Google Scholar] [CrossRef]
- Ayanoğlu, F.; Kaya, D.A.; Koçer, O. Promising bay laurel (Laurus nobilis L.) genotypes for fruit production. Int. J. Chem. Technol. 2018, 2, 161–167. [Google Scholar] [CrossRef]
- Ibdah, M.; Muchlinski, A.; Yahyaa, M.; Nawade, B.; Tholl, D. Carrot volatile terpene metabolism: Terpene diversity and biosynthetic genes. In The Carrot Genome; Springer: Berlin/Heidelberg, Germany, 2019; pp. 279–293. [Google Scholar]
- Vaughn, S.F. Plant Volatiles; eLS: Chicago, IL, USA, 2001. [Google Scholar]
- Dudareva, N.; Pichersky, E. Metabolic engineering of plant volatiles. Curr. Opin. Biotechnol. 2008, 19, 181–189. [Google Scholar] [CrossRef]
- Nawade, B.; Yahyaa, M.; Reuveny, H.; Shaltiel-Harpaz, L.; Eisenbach, O.; Faigenboim, A.; Bar-Yaakov, I.; Holland, D.; Ibdah, M. Profiling of volatile terpenes from almond (Prunus dulcis) young fruits and characterization of seven terpene synthase genes. Plant Sci. 2019, 287, 110187. [Google Scholar] [CrossRef]
- Goff, S.A.; Klee, H.J. Plant volatile compounds: Sensory cues for health and nutritional value? Science 2006, 311, 815–819. [Google Scholar] [CrossRef]
- Singletary, K. Bay leaf: Potential health benefits. Nutr. Today 2021, 56, 202–208. [Google Scholar] [CrossRef]
- Nawade, B.; Yahyaa, M.; Lewinsohn, E.; Ibdah, M. Vegetative and fruit volatiles for human consumption. In Biology of Plant Volatiles; CRC Press: Boca Raton, FL, USA, 2020; pp. 79–95. [Google Scholar]
- Wilson, L. Spices and flavoring crops: Leaf and floral structures. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; pp. 84–92. [Google Scholar]
- Fasseas, C.; Akoumianaki-Ioannidou, A. Some anatomical features of Laurus nobilis leaves contributing to its suitability as ornamental plant in temperate climates. In Proceedings of the I International Symposium on Woody Ornamentals of the Temperate Zone 885, Pruhonice, Czech Republic, 26 May 2008; pp. 99–108. [Google Scholar]
- Kondraskov, P.; Schütz, N.; Schüßler, C.; de Sequeira, M.M.; Guerra, A.S.; Caujapé-Castells, J.; Jaén-Molina, R.; Marrero-Rodríguez, Á.; Koch, M.A.; Linder, P. Biogeography of mediterranean hotspot biodiversity: Re-evaluating the ‘tertiary relict’ hypothesis of macaronesian laurel forests. PLoS ONE 2015, 10, e0132091. [Google Scholar] [CrossRef]
- Rodríguez-Sánchez, F.; Guzmán, B.; Valido, A.; Vargas, P.; Arroyo, J. Late Neogene history of the laurel tree (Laurus L., Lauraceae) based on phylogeographical analyses of Mediterranean and Macaronesian populations. J. Biogeogr. 2009, 36, 1270–1281. [Google Scholar] [CrossRef] [Green Version]
- Huxley, A.; Griffiths, M.; Levy, M. Dictionary of Gardening: The New Royal Horticultural Society; Macmillan Press: London, UK; New York, NY, USA, 1992. [Google Scholar]
- Huxley, A.J.; Griffiths, M. New Royal Horticultural Society Dictionary of Gardening; Grove’s Dictionaries Inc.: New York, NY, USA, 1999. [Google Scholar]
- Taylor, J. The Milder Garden; Orion Publishing Group: London, UK, 1990. [Google Scholar]
- Souayah, N.; Khouja, M.L.; Khaldi, A.; Rejeb, M.N.; Bouzid, S. Breeding improvement of Laurus nobilis L. by conventional and in vitro propagation techniques. J. Herbs Spices. Med. Plants 2002, 9, 101–105. [Google Scholar] [CrossRef]
- Takos, I.A. Seed dormancy in bay laurel (Laurus nobilis L.). New For. 2001, 21, 105–114. [Google Scholar] [CrossRef]
- Tilki, F. Influence of pretreatment and desiccation on the germination of Laurus nobilis L. seeds. J. Environ. Biol. 2004, 25, 157–161. [Google Scholar] [PubMed]
- Konstantinidou, E.; Takos, I.; Merou, T. Desiccation and storage behavior of bay laurel (Laurus nobilis L.) seeds. Eur. J. For. Res. 2008, 127, 125–131. [Google Scholar] [CrossRef]
- Ambrose, D.C.; Manickavasagan, A.; Naik, R. Leafy Medicinal Herbs: Botany, Chemistry, Postharvest Technology and Uses; CABI: London, UK, 2016.
- Kumar, S.; Singh, J.; Sharma, A. Bay leaves. In Handbook of Herbs and Spices; Elsevier: Amsterdam, The Netherlands, 2001; pp. 52–61. [Google Scholar]
- Al Gabbiesh, A.H.; Ghabeish, M.; Kleinwächter, M.; Selmar, D. Plant regeneration through somatic embryogenesis from calli derived from leaf bases of Laurus nobilis L. (Lauraceae). Plant Tissue Cult. Biotechnol. 2014, 24, 213–221. [Google Scholar] [CrossRef]
- Chourfi, A.; Alaoui, T.; Echchgadda, G. In vitro propagation of the bay laurel (Laurus nobilis L.) in Morocco. SAJEB 2014, 4, 96–103. [Google Scholar] [CrossRef]
- Foqué, D.; Braekman, P.; Pieters, J.G.; Nuyttens, D. A vertical spray boom application technique for conical bay laurel (Laurus nobilis) plants. Crop Prot. 2012, 41, 113–121. [Google Scholar] [CrossRef]
- Zeity, M. First Record of the Bay Sucker Trioza Alacris Flor (Triozidae: Hemiptera) in Syria; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2018. [Google Scholar]
- Vitale, A.; Polizzi, G. Occurrence of Pestalotiopsis uvicola causing leaf spots and stem blight on bay laurel (Laurus nobilis) in Sicily. Plant Dis. 2005, 89, 1362. [Google Scholar] [CrossRef]
- Duarte, A.D.F.; Andreazza, F.; Nava, D.E.; Da Cunha, U.S. Polyphagotarsonemus latus (Trombidiformes: Tarsonemidae) on Laurus nobilis (Polycarpicae: Lauraceae): Report of infestation and damage. Syst. Appl. Acarol. 2021, 26, 1614–1618. [Google Scholar] [CrossRef]
- Weiss, E.A. Spice Crops; CABI: London, UK, 2002.
- Akgül, A. Spice Science and Technology; Turkish Association of Food Technologists Publ. No. 15: Ankara, Turkey, 1993. [Google Scholar]
- Raghavan, S. Handbook of Spices, Seasonings, and Flavorings; CRC: Boca Raton, FL, USA, 2007. [Google Scholar]
- Nadeem, M.A.; Aasim, M.; Kırıcı, S.; Karık, Ü.; Nawaz, M.A.; Yılmaz, A.; Maral, H.; Khawar, K.M.; Baloch, F.S. Laurel (Laurus nobilis L.): A less-known medicinal plant to the world with diffusion, genomics, phenomics, and metabolomics for genetic improvement. In Biotechnological Approaches for Medicinal and Aromatic Plants; Springer: Berlin/Heidelberg, Germany, 2018; pp. 631–653. [Google Scholar]
- Ayanoğlu, F.; Kaya, D.; Mert, A.; Köse, E. Determination of quality aspects and selection of native grown laurel (Laurus nobilis L.) in Hatay province of Turkey. In Proceedings of the The First Mediterranean Symposium on Medicinal and Aromatic Plants (MESMAP), Famagusta, Cyprus, 17–20 April 2013; pp. 17–20. [Google Scholar]
- Bulut, M.Ç.; Özmen, C.Y.; Ergül, A.; Ayanoğlu, F. Genetic characterization of Bay Laurel (Laurus nobilis L.) populations using microsatellite markers and flow cytometry. Mustafa Kemal Üniversitesi Ziraat Fakültesi Dergisi 2018, 23, 242–253. [Google Scholar]
- Bolarić, S.; Dunkić, V.D.; Bezić, N.; Kremer, D.; Karlović, K.; Tuković, A.; Jelak, L.; Srečec, S. Influence of geographic position, leaf surface and genetic variability on content of total essential oils in 12 distinct populations of Bay Laurel (Laurus nobilis L.). Agric. Conspec. Sci. 2019, 84, 83–89. [Google Scholar]
- Choudhary, D.; Kala, S.; Todaria, N.; Dasgupta, S.; Kinhal, G.; Kollmair, M. Essential oil from Bay leaves in India and Nepal: An analysis for quality oriented value chain development. Int. J. Med. Aromat. Plants 2013, 3, 11–17. [Google Scholar]
- Southwell, I.A.; Russell, M.F.; Maddox, C.D.; Wheeler, G.S. Differential metabolism of 1,8-cineole in insects. J. Chem. Ecol. 2003, 29, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Franks, S.J.; Wheeler, G.S.; Goodnight, C. Genetic variation and evolution of secondary compounds in native and introduced populations of the invasive plant Melaleuca quinquenervia. Evol. Int. J. Org. Evol. 2012, 66, 1398–1412. [Google Scholar] [CrossRef]
- Mercier, B.; Prost, J.; Prost, M. The essential oil of turpentine and its major volatile fraction (α-and β-pinenes): A review. Int. J. Occup. Med. Environ. Health 2009, 22, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.H.; Nishida, R. Methyl eugenol: Its occurrence, distribution, and role in nature, especially in relation to insect behavior and pollination. J. Insect Sci. 2012, 12, 56. [Google Scholar] [CrossRef] [Green Version]
- Pino, J.; Borges, P.; Roncal, E. The chemical composition of laurel leaf oil from various origins. Mol. Nutr. Food Res. 1993, 37, 592–595. [Google Scholar] [CrossRef]
- Marzouki, H.; Khaldi, A.; Chamli, R.; Bouzid, S.; Piras, A.; Falconieri, D.; Marongiu, B. Biological activity evaluation of the oils from Laurus nobilis of Tunisia and Algeria extracted by supercritical carbon dioxide. Nat. Prod. Res. 2009, 23, 230–237. [Google Scholar] [CrossRef]
- Jemâa, J.M.B.; Tersim, N.; Toudert, K.T.; Khouja, M.L. Insecticidal activities of essential oils from leaves of Laurus nobilis L. from Tunisia, Algeria and Morocco, and comparative chemical composition. J. Stored Prod. Res. 2012, 48, 97–104. [Google Scholar] [CrossRef]
- Lira, P.D.L.; Retta, D.; Tkacik, E.; Ringuelet, J.; Coussio, J.; Van Baren, C.; Bandoni, A. Essential oil and by-products of distillation of bay leaves (Laurus nobilis L.) from Argentina. Ind. Crops Prod. 2009, 30, 259–264. [Google Scholar] [CrossRef]
- Řebíčková, K.; Bajer, T.; Šilha, D.; Ventura, K.; Bajerová, P. Comparison of chemical composition and biological properties of essential oils obtained by hydrodistillation and steam distillation of Laurus nobilis L. Plant Foods Hum. Nutr. 2020, 75, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Fidan, H.; Stefanova, G.; Kostova, I.; Stankov, S.; Damyanova, S.; Stoyanova, A.; Zheljazkov, V.D. Chemical composition and antimicrobial activityof Laurus nobilis L. essential oils from Bulgaria. Molecules 2019, 24, 804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Politeo, O.; Jukić, M.; Miloš, M. Chemical composition and antioxidant activity of free volatile aglycones from laurel (Laurus nobilis L.) compared to its essential oil. Croat. Chem. Acta. 2007, 80, 121–126. [Google Scholar]
- Fiorini, C.; Fouraste, I.; David, B.; Bessiere, J. Composition of the flower, leaf and stem essential oils from Laurus nobilis. Flavour Fragr. J. 1997, 12, 91–93. [Google Scholar] [CrossRef]
- Braun, N.A.; Meier, M.; Kohlenberg, B.; Hammerschmidt, F.-J. δ-Terpinyl acetate. A new natural component from the essential leaf oil of Laurus nobilis L. (Lauraceae). J. Essent. Oil Res. 2001, 13, 95–97. [Google Scholar] [CrossRef]
- Hassiotis, C.N.; Dina, E.I. The effects of laurel (Laurus nobilis L.) on development of two mycorrhizal fungi. Int. Biodeterior. Biodegradation 2011, 65, 628–634. [Google Scholar] [CrossRef]
- Stefanova, G.; Girova, T.; Gochev, V.; Stoyanova, M.; Petkova, Z.; Stoyanova, A.; Zheljazkov, V.D. Comparative study on the chemical composition of laurel (Laurus nobilis L.) leaves from Greece and Georgia and the antibacterial activity of their essential oil. Heliyon 2020, 6, e05491. [Google Scholar] [CrossRef]
- Kaur, M.; Chahal, K.; Kumar, A.; Kaur, R. Nematicidal activity of bay leaf (Laurus nobilis L.) essential oil and its components against Meloidogyne incognita. J. Entomol. Zool. Stud. 2018, 6, 1057–1064. [Google Scholar]
- Hadjibagher Kandi, M.N.; Sefidkon, F. The influense of drying methods on essential oil content and composition of Laurus nobilis L. J. Essent. Oil-Bear. Plants 2011, 14, 302–308. [Google Scholar] [CrossRef]
- Moghtader, M.; Salari, H. Comparative survey on the essential oil composition from the leaves and flowers of Laurus nobilis L. from Kerman province. J. Ecol. Nat. 2012, 4, 150–153. [Google Scholar] [CrossRef]
- Verdian-Rizi, M. Chemical composition and larvicidal activity of the essential oil of Laurus nobilis L. from Iran. Iran. J. Pharm. Sci. 2009, 5, 47–50. [Google Scholar]
- Shokoohinia, Y.; Yegdaneh, A.; Amin, G.; Ghannadi, A. Seasonal variations of Laurus nobilis L. leaves volatile oil components in Isfahan, Iran. Res. J. Pharmacogn. 2014, 1, 1–6. [Google Scholar]
- Amin, G.; Sourmaghi, M.S.; Jaafari, S.; Hadjagaee, R.; Yazdinezhad, A. Influence of phenological stages and method of distillation on Iranian cultivated bay leaves volatile oil. Pak. J. Biol. Sci. 2007, 10, 2895–2899. [Google Scholar] [CrossRef] [Green Version]
- Caredda, A.; Marongiu, B.; Porcedda, S.; Soro, C. Supercritical carbon dioxide extraction and characterization of Laurus nobilis essential oil. J. Agric. Food Chem. 2002, 50, 1492–1496. [Google Scholar] [CrossRef]
- Saab, A.M.; Tundis, R.; Loizzo, M.R.; Lampronti, I.; Borgatti, M.; Gambari, R.; Menichini, F.; Esseily, F.; Menichini, F. Antioxidant and antiproliferative activity of Laurus nobilis L. (Lauraceae) leaves and seeds essential oils against K562 human chronic myelogenous leukaemia cells. Nat. Prod. Res. 2012, 26, 1741–1745. [Google Scholar] [CrossRef]
- Flamini, G.; Tebano, M.; Cioni, P.L.; Ceccarini, L.; Ricci, A.S.; Longo, I. Comparison between the conventional method of extraction of essential oil of Laurus nobilis L. and a novel method which uses microwaves applied in situ, without resorting to an oven. J. Chromatogr. A 2007, 1143, 36–40. [Google Scholar] [CrossRef]
- Abu-Dahab, R.; Kasabri, V.; Afifi, F.U. Evaluation of the volatile oil composition and antiproliferative activity of Laurus nobilis L. (Lauraceae) on breast cancer cell line models. Rec. Nat. Prod. 2014, 8, 136. [Google Scholar]
- Dahak, K.; Bouamama, H.; Benkhalti, F.; Taourirte, M. Drying methods and their implication on quality, quantity and antimicrobial activity of the essential oil of Laurus nobilis L. from Morocco. Online J. Biol. Sci. 2014, 14, 94–101. [Google Scholar] [CrossRef] [Green Version]
- Derwich, E.; Benziane, Z.; Boukir, A. Chemical composition and antibacterial activity of leaves essential oil of Laurus nobilis from Morocco. AJBAS 2009, 3, 3818–3824. [Google Scholar]
- Ramos, C.; Teixeira, B.; Batista, I.; Matos, O.; Serrano, C.; Neng, N.; Nogueira, J.; Nunes, M.; Marques, A. Antioxidant and antibacterial activity of essential oil and extracts of bay laurel Laurus nobilis Linnaeus (Lauraceae) from Portugal. Nat. Prod. Res. 2012, 26, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Maroto, M.C.; Pérez-Coello, M.S.; Cabezudo, M.D. Effect of drying method on the volatiles in bay leaf (Laurus nobilis L.). J. Agric. Food Chem. 2002, 50, 4520–4524. [Google Scholar] [CrossRef] [PubMed]
- Zeković, Z.P.; Lepojević, Ž.D.; Mujić, I.O. Laurel extracts obtained by steam distillation, supercritical fluid and solvent extraction. J. Nat. Prod. 2009, 2, 104–109. [Google Scholar]
- Simić, A.; Soković, M.; Ristić, M.; Grujić-Jovanović, S.; Vukojević, J.; Marin, P. The chemical composition of some Lauraceae essential oils and their antifungal activities. Phytother. Res. 2004, 18, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Ivanović, J.; Mišić, D.; Ristić, M.; Pešić, O.; Žižović, I. Supercritical CO2 extract and essential oil of bay (Laurus nobilis L.): Chemical composition and antibacterial activity. J. Serb. Chem. Soc. 2010, 75, 395–404. [Google Scholar] [CrossRef]
- Riabov, P.A.; Micić, D.; Božović, R.B.; Jovanović, D.V.; Tomić, A.; Šovljanski, O.; Filip, S.; Tosti, T.; Ostojić, S.; Blagojević, S. The chemical, biological and thermal characteristics and gastronomical perspectives of Laurus nobilis essential oil from different geographical origin. Ind. Crops Prod. 2020, 151, 112498. [Google Scholar] [CrossRef]
- Hagibaghercandi, M.; Sefidkon, F.; Poorherave, M.; Mirza, M. Extraction, identification and comparison of chemical composition of the stem, leaf and flower essential oil from Laurus nobilis L. IJMAPR 2009, 25, 216–227. [Google Scholar]
- Marzouki, H.; Piras, A.; Marongiu, B.; Rosa, A.; Dessi, M.A. Extraction and separation of volatile and fixed oils from berries of Laurus nobilis L. by supercritical CO2. Molecules 2008, 13, 1702–1711. [Google Scholar] [CrossRef] [Green Version]
- Flamini, G.; Cioni, P.L.; Morelli, I. Differences in the fragrances of pollen and different floral parts of male and female flowers of Laurus nobilis. J. Agric. Food Chem. 2002, 50, 4647–4652. [Google Scholar] [CrossRef]
- Mssillou, I.; Agour, A.; El Ghouizi, A.; Hamamouch, N.; Lyoussi, B.; Derwich, E. Chemical composition, antioxidant activity, and antifungal effects of essential oil from Laurus nobilis L. flowers growing in Morocco. J. Food Qual. 2020, 2020, 8819311. [Google Scholar] [CrossRef]
- Yahyaa, M.; Matsuba, Y.; Brandt, W.; Doron-Faigenboim, A.; Bar, E.; McClain, A.; Davidovich-Rikanati, R.; Lewinsohn, E.; Pichersky, E.; Ibdah, M. Identification, functional characterization, and evolution of terpene synthases from a basal dicot. Plant Physiol. 2015, 169, 1683–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafizoǧlu, H.; Reunanen, M. Studies on the components of Lauras nobilis from Turkey with special reference to Laurel berry fat. Lipid/Fett 1993, 95, 304–308. [Google Scholar] [CrossRef]
- Castilho, P.C.; do Céu Costa, M.; Rodrigues, A.; Partidário, A. Characterization of laurel fruit oil from Madeira Island, Portugal. J. Am. Oil Chem. Soc. 2005, 82, 863–868. [Google Scholar] [CrossRef]
- Marzouki, H.; Piras, A.; Salah, K.B.H.; Medini, H.; Pivetta, T.; Bouzid, S.; Marongiu, B.; Falconieri, D. Essential oil composition and variability of Laurus nobilis L. growing in Tunisia, comparison and chemometric investigation of different plant organs. Nat. Prod. Res. 2009, 23, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Boulila, A.; Hassen, I.; Haouari, L.; Mejri, F.; Amor, I.B.; Casabianca, H.; Hosni, K. Enzyme-assisted extraction of bioactive compounds from bay leaves (Laurus nobilis L.). Ind. Crops Prod. 2015, 74, 485–493. [Google Scholar] [CrossRef]
- Ozek, T.; Bozan, B.; Baser, K.H. Supercritical CO2 extraction of volatile components from leaves of Laurus nobilis L. Chem. Nat. Compd. 1998, 34, 668–671. [Google Scholar] [CrossRef]
- Bendjersi, F.Z.; Tazerouti, F.; Belkhelfa-Slimani, R.; Djerdjouri, B.; Meklati, B.Y. Phytochemical composition of the Algerian Laurus nobilis L. Leaves extracts obtained by solvent-free microwave extraction and investigation of their antioxidant activity. J. Essent. Oil Res. 2016, 28, 202–210. [Google Scholar] [CrossRef]
- Tunç, M.T.; Koca, İ. Optimization of ohmic heating assisted hydrodistillation of cinnamon and bay leaf essential oil. J. Food Process Eng. 2021, 44, e13635. [Google Scholar] [CrossRef]
- Sellami, I.H.; Wannes, W.A.; Bettaieb, I.; Berrima, S.; Chahed, T.; Marzouk, B.; Limam, F. Qualitative and quantitative changes in the essential oil of Laurus nobilis L. leaves as affected by different drying methods. Food Chem. 2011, 126, 691–697. [Google Scholar] [CrossRef]
- Asekun, O.; Grierson, D.; Afolayan, A.J. Effects of drying methods on the quality and quantity of the essential oil of Mentha longifolia L. subsp. Capensis. Food Chem. 2007, 101, 995–998. [Google Scholar] [CrossRef]
- Arroyo-García, R.; Martínez-Zapater, J.M.; Prieto, J.F.; Álvarez-Arbesú, R. AFLP evaluation of genetic similarity among laurel populations (Laurus L.). Euphytica 2001, 122, 155–164. [Google Scholar] [CrossRef]
- Mohamed, A.-S.H.; Ahmed, W.; Rabia, S.S.; Mourad, M.M. Implications of morphology and molecular criteria in taxonomy of lauraceae juss. Egypt. J. Exp. Biol. (Bot.) 2016, 12, 45–52. [Google Scholar]
- Sevindik, E. Molecular genetic diversity of some Laurus nobilis L. (Lauraceae) populations grown in the Aegean region/Turkey. Adv. Food Sci. 2019, 41, 28–31. [Google Scholar]
- Powell, W.; Machray, G.C.; Provan, J. Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1996, 1, 215–222. [Google Scholar] [CrossRef]
- Marzouki, H.; Nasri, N.; Jouaud, B.; Bonnet, C.; Khaldi, A.; Bouzid, S.; Fady, B. Population genetic structure of Laurus nobilis L. inferred from transferred nuclear microsatellites. Silvae Genet. 2009, 58, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, J.M.; Rigueiro, C.; Rodríguez, R.; Hampe, A.; Valido, A.; Rodríguez-Sánchez, F.; Jordano, P. Isolation and characterization of 20 microsatellite loci for laurel species (Laurus, Lauraceae). Am. J. Bot. 2010, 97, e26–e30. [Google Scholar] [CrossRef] [PubMed]
- Trindade, H. Molecular biology of aromatic plants and spices. A review. Flavour Fragr. J. 2010, 25, 272–281. [Google Scholar] [CrossRef]
- Pacini, E.; Sciannandrone, N.; Nepi, M. Floral biology of the dioecious species Laurus nobilis L. (Lauraceae). Flora Morphol. Distrib. Funct. Ecol. Plants 2014, 209, 153–163. [Google Scholar] [CrossRef]
- Shahsavari, S. An overview of the most important medicinal plants used in iranian traditional medicine for the treatment of kidney stones: A mini-review article. Plant Biotechnol. Persa 2021, 3, 1–4. [Google Scholar] [CrossRef]
- Aytürk, Ö.; Meral, Ü. Structural analysis of reproductive development in staminate flowers of Laurus nobilis L. Not. Sci. Biol. 2012, 4, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Royandazagh, D. Potential of flow cytometry in sex determination and in vitro micropropagation of Laurus nobilis L. Appl. Ecol. Environ. Res. 2019, 17, 5953–5964. [Google Scholar] [CrossRef]
- Heikrujam, M.; Sharma, K.; Prasad, M.; Agrawal, V. Review on different mechanisms of sex determination and sex-linked molecular markers in dioecious crops: A current update. Euphytica 2015, 201, 161–194. [Google Scholar] [CrossRef]
- Chahal, K.; Kaur, M.; Bhardwaj, U.; Singla, N.; Kaur, A.; Kaur, M.; Bhardwaj, U.; Singla, N.; Kaur, A. A review on chemistry and biological activities of Laurus nobilis L. essential oil. J. Pharmacogn. Phytochem. 2017, 6, 1153–1161. [Google Scholar]
- Patrakar, R.; Mansuriya, M.; Patil, P. Phytochemical and pharmacological review on Laurus nobilis. Int. J. Pharm. Chem. 2012, 1, 595–602. [Google Scholar]
- Dammak, I.; Hamdi, Z.; El Euch, S.K.; Zemni, H.; Mliki, A.; Hassouna, M.; Lasram, S. Evaluation of antifungal and anti-ochratoxigenic activities of Salvia officinalis, Lavandula dentata and Laurus nobilis essential oils and a major monoterpene constituent 1, 8-cineole against Aspergillus carbonarius. Ind. Crops Prod. 2019, 128, 85–93. [Google Scholar] [CrossRef]
- Olmedo, R.H.; Asensio, C.M.; Grosso, N.R. Thermal stability and antioxidant activity of essential oils from aromatic plants farmed in Argentina. Ind. Crops Prod. 2015, 69, 21–28. [Google Scholar] [CrossRef]
- Basak, S.S.; Candan, F. Effect of Laurus nobilis L. essential oil and its main components on α-glucosidase and reactive oxygen species scavenging activity. Iran. J. Pharm. Res. 2013, 12, 367. [Google Scholar]
- Nafis, A.; Kasrati, A.; Jamali, C.A.; Custódio, L.; Vitalini, S.; Iriti, M.; Hassani, L. A comparative study of the in vitro antimicrobial and synergistic effect of essential oils from Laurus nobilis L. and Prunus armeniaca L. from Morocco with antimicrobial drugs: New approach for health promoting products. Antibiotics 2020, 9, 140. [Google Scholar] [CrossRef] [Green Version]
- Tomar, O.; Akarca, G.; Gök, V.; Ramadan, M.F. Composition and antibacterial effects of Laurel (Laurus nobilis L.) leaves essential oil. J. Essent. Oil-Bear. Plants 2020, 23, 414–421. [Google Scholar] [CrossRef]
- Nenadis, N.; Papapostolou, M.; Tsimidou, M.Z. Suggestions on the contribution of methyl eugenol and eugenol to bay laurel (Laurus nobilis L.) essential oil preservative activity through radical scavenging. Molecules 2021, 26, 2342. [Google Scholar] [CrossRef]
- Yilmaz, E.S.; Timur, M.; Aslim, B. Antimicrobial, antioxidant activity of the essential oil of Bay Laurel from Hatay, Turkey. J. Essent. Oil-Bear. Plants 2013, 16, 108–116. [Google Scholar] [CrossRef]
- Taoudiat, A.; Djenane, D.; Ferhat, Z.; Spigno, G. The effect of Laurus nobilis L. essential oil and different packaging systems on the photo-oxidative stability of Chemlal extra-virgin olive oil. J. Food Sci. Technol. 2018, 55, 4212–4222. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, S.; Lloria, R.; Jaime, L.; Ibanez, E.; Senorans, F.; Reglero, G. Supercritical fluid extraction of antioxidant and antimicrobial compounds from Laurus nobilis L. Chemical and functional characterization. Eur. Food Res. Technol. 2006, 222, 565–571. [Google Scholar] [CrossRef]
- Dall’Acqua, S.; Cervellati, R.; Speroni, E.; Costa, S.; Guerra, M.C.; Stella, L.; Greco, E.; Innocenti, G. Phytochemical composition and antioxidant activity of Laurus nobilis L. leaf infusion. J. Med. Food 2009, 12, 869–876. [Google Scholar] [CrossRef] [PubMed]
- Bozan, B.; Karakaplan, U. Antioxidants from laurel (Laurus nobilis L.) berries: Influence of extraction procedure on yield and antioxidant activity of extracts. Acta Aliment. 2007, 36, 321–328. [Google Scholar] [CrossRef]
- Speroni, E.; Cervellati, R.; Dall’Acqua, S.; Guerra, M.C.; Greco, E.; Govoni, P.; Innocenti, G. Gastroprotective effect and antioxidant properties of different Laurus nobilis L. leaf extracts. J. Med. Food 2011, 14, 499–504. [Google Scholar] [CrossRef]
- Ozcan, B.; Esen, M.; Sangun, M.K.; Coleri, A.; Caliskan, M. Effective antibacterial and antioxidant properties of methanolic extract of Laurus nobilis seed oil. J. Environ. Biol. 2010, 31, 637–641. [Google Scholar]
- Kaurinovic, B.; Popovic, M.; Vlaisavljevic, S. In vitro and in vivo effects of Laurus nobilis L. leaf extracts. Molecules 2010, 15, 3378–3390. [Google Scholar] [CrossRef] [Green Version]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods-a review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Nabila, B.; Piras, A.; Fouzia, B.; Falconieri, D.; Kheira, G.; Fedoul, F.-F.; Majda, S.-R. Chemical composition and antibacterial activity of the essential oil of Laurus nobilis leaves. Nat. Prod. Res. 2022, 36, 989–993. [Google Scholar] [CrossRef]
- Caputo, L.; Nazzaro, F.; Souza, L.F.; Aliberti, L.; De Martino, L.; Fratianni, F.; Coppola, R.; De Feo, V. Laurus nobilis: Composition of essential oil and its biological activities. Molecules 2017, 22, 930. [Google Scholar] [CrossRef] [PubMed]
- Bayar, Y.; Onaran, A.; Yilar, M.; Gul, F. Determination of the essential oil composition and the antifungal activities of bilberry (Vaccinium myrtillus L.) and bay laurel (Laurus nobilis L.). J. Essent. Oil-Bear. Plants 2018, 21, 548–555. [Google Scholar] [CrossRef]
- Xu, S.; Yan, F.; Ni, Z.; Chen, Q.; Zhang, H.; Zheng, X. In vitro and in vivo control of Alternaria alternata in cherry tomato by essential oil from Laurus nobilis of Chinese origin. J. Sci. Food Agric. 2014, 94, 1403–1408. [Google Scholar] [CrossRef] [PubMed]
- Moumni, M.; Romanazzi, G.; Najar, B.; Pistelli, L.; Ben Amara, H.; Mezrioui, K.; Karous, O.; Chaieb, I.; Allagui, M.B. Antifungal activity and chemical composition of seven essential oils to control the main seedborne fungi of cucurbits. Antibiotics 2021, 10, 104. [Google Scholar] [CrossRef] [PubMed]
- Belasli, A.; Ben Miri, Y.; Aboudaou, M.; Aït Ouahioune, L.; Montañes, L.; Ariño, A.; Djenane, D. Antifungal, antitoxigenic, and antioxidant activities of the essential oil from laurel (Laurus nobilis L.): Potential use as wheat preservative. Food Sci. Nutr. 2020, 8, 4717–4729. [Google Scholar] [CrossRef]
- Peixoto, L.R.; Rosalen, P.L.; Ferreira, G.L.S.; Freires, I.A.; de Carvalho, F.G.; Castellano, L.R.; de Castro, R.D. Antifungal activity, mode of action and anti-biofilm effects of Laurus nobilis Linnaeus essential oil against Candida spp. Arch. Oral Biol. 2017, 73, 179–185. [Google Scholar] [CrossRef]
- Cherrat, L.; Espina, L.; Bakkali, M.; García-Gonzalo, D.; Pagán, R.; Laglaoui, A. Chemical composition and antioxidant properties of Laurus nobilis L. and Myrtus communis L. essential oils from Morocco and evaluation of their antimicrobial activity acting alone or in combined processes for food preservation. J. Sci. Food Agric. 2014, 94, 1197–1204. [Google Scholar] [CrossRef]
- da Silveira, S.M.; Luciano, F.B.; Fronza, N.; Cunha Jr, A.; Scheuermann, G.N.; Vieira, C.R.W. Chemical composition and antibacterial activity of Laurus nobilis essential oil towards foodborne pathogens and its application in fresh Tuscan sausage stored at 7 C. LWT-Food Sci. Technol. 2014, 59, 86–93. [Google Scholar] [CrossRef]
- De Corato, U.; Maccioni, O.; Trupo, M.; Di Sanzo, G. Use of essential oil of Laurus nobilis obtained by means of a supercritical carbon dioxide technique against post harvest spoilage fungi. Crop Prot. 2010, 29, 142–147. [Google Scholar] [CrossRef]
- D’Amato, S.; Serio, A.; López, C.C.; Paparella, A. Hydrosols: Biological activity and potential as antimicrobials for food applications. Food Control 2018, 86, 126–137. [Google Scholar] [CrossRef]
- Haouas, D. Natural mediterranean plants products: A sustainable approach for integrated insect pests management. In Agriculture Productivity in Tunisia under Stressed Environment; Springer: Berlin/Heidelberg, Germany, 2021; pp. 137–154. [Google Scholar]
- Nawade, B.; Shaltiel-Harpaz, L.; Yahyaa, M.; Kabaha, A.; Kedoshim, R.; Bosamia, T.C.; Ibdah, M. Characterization of terpene synthase genes potentially involved in black fig fly (Silba adipata) interactions with Ficus carica. Plant Sci. 2020, 298, 110549. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [Green Version]
- Salehi, T.; Karimi, J.; Hasanshahi, G.; Askarianzadeh, A.; Abbasipour, H.; Garjan, A.S. The effect of essential oils from Laurus nobilis and Myrtus commonis on the adults of mediterranean flour moth, Ephestia kuehniella Zeller (Lep.: Pyralidae). J. Essent. Oil-Bear. Plants 2014, 17, 553–561. [Google Scholar] [CrossRef]
- Damiani, N.; Fernández, N.J.; Porrini, M.P.; Gende, L.B.; Álvarez, E.; Buffa, F.; Brasesco, C.; Maggi, M.D.; Marcangeli, J.A.; Eguaras, M.J. Laurel leaf extracts for honeybee pest and disease management: Antimicrobial, microsporicidal, and acaricidal activity. Parasitol. Res. 2014, 113, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, M.; Safaralizade, M.H.; Valizadegan, O.; Amin, B.H.H. Efficacy of three plant essential oils, Azadirachta indica (Adr. Juss.), Eucalyptus camaldulensis (Dehn.) and Laurus nobilis (L.) on mortality cotton aphids, Aphis gossypii Glover (Hem: Aphididae). Arch. Phytopathol. Plant Prot. 2013, 46, 1093–1101. [Google Scholar] [CrossRef]
- Soltani, A.; Tassnim, D.; Mediouni-Ben Jemâa, J. Fumigant potential of essential oils from Laurus nobilis (Lau-raceae) against the date moth Ectomyelois ceratoniae (Pyralidae). In Proceedings of the Proceedings of the 1st International Electronic Conference on Entomology, online, 1–15 July 2021; pp. 1–15. [Google Scholar]
- Shaltiel, L.; Coll, M. Reduction of pear psylla damage by the predatory bug Anthocoris nemoralis (Heteroptera: Anthocoridae): The importance of orchard colonization time and neighboring vegetation. Biocontrol Sci. Technol. 2004, 14, 811–821. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cultivar Name | Main Features | Common Name |
|---|---|---|
| Angustifolia | Blades much narrower, usually lanceolate to oblanceolate. Leaves are a lighter or brighter green than typical species. | Willow-leaved laural |
| Aurea/Aureum | Golden yellow foliage, brightest in winter and spring, and dense clusters of small, greenish-yellow male flowers in spring | Golden bay |
| Baccalia/Bacalia | In old literature, Bacalia was mentioned as the commonest laurel and the one that bears the greatest number of berries. | |
| Bay junior | Compact and slower growth habitat grows more slowly with a smaller leaf but produces very full, dense foliage-medium-wide elliptic leaves. | |
| Borms | The cold-resistant cultivar produces fewer or no flowers | |
| Borziana var. borzianaBég | Elongated, narrower leaf with a thinner blade; fruits more spherical large to ca. 1/2 inch. | Borzis bay |
| Brilliant times | Stem is often reddish in new shoots, contrasting and showy at times. Leaves are bright yellow, faintly tinged red on margins at young later yellowish-green. | |
| Celtic queen | Fast-growing—can grow to 8 feet tall in 3 years and is hardy tolerating as low as −4 F, deep green foliage | |
| Chichester | Mottled and sectored leaf with narrow elliptic to linear-elliptic blade much as ‘Angustifolia’ with chimera | |
| Crispa/Undulata | Leaf margin distinctly undulate, egg-shaped fruits, low shrub seldom higher than 4 to 6 feet. | Wavy-leaved bay |
| Cylindrocarpa | Leaves are fairly wide and ovate; cylindrical fruits. | Cylinder-fruited bay |
| Cyprian | Shorter in height, darker, blackish-green leaves and their margins are crisped and imbricated. | |
| Delphic | Leaves are uniformly green, and fruits are green-tinged red with large sizes. | |
| Emerald Wave@Monem | Wavy-edged, emerald green leaves; slender, upright form grows slowly and gracefully into a small pyramidal tree, ideal for topiary. | |
| Eriobotryfolia | Very large, wide, toothed leaves resemble Eriobotrya. | Loquat-leaved bay |
| Feys | Cold hardy cultivar. | |
| Flore Pleno | A double-flowered version of the broad-leaved Dutch myrtle. | Double-flowered bay |
| Glauca | More glaucous or bluish (waxy) leaves. | Glaucous-leaved bay |
| Goldstein | Slightly undulate classic bay leaf with white midrib. | |
| Grandiflora | Larger fruit diameter than average. | |
| Hex brilliant | Leaves irregularly margined white to cream and with a thin margin, irregularly twisted and very asymmetrical. | |
| Holy land | Origin–Israel, leaves are slightly wavy on the edges. | |
| Latifolia | Leaves are broad and smooth, less hardy than most other varieties, native to Spain, Italy, and Asia. | Broad-leaved bay |
| Ligustrifolia | Privet-shaped leaves. | Privet-leaved bay |
| Little Ragu® ‘MonRik’ | A more compact sweet bay with highly aromatic, deep green foliage that emerges on unique red-tinged stems. Fabulous fragrance. | |
| Macrocarpa | Much larger fruit diameter. | Large-fruited bay |
| Macroclada | Tall plants, leaves are rounded apex, tapering to base, stout while fruits are globular, large, ca. 1/2 inch. | Large bay |
| Microcarpa | Small fruit diameter. | |
| Moonspot | Leaves mottled and sectored white to 35%. | |
| Multiflora | Flowers in multiples or numerous. | Many-flowered bay |
| Mustax | Very large blade and the blades are more flaccid. Leaves are a whitish hue. It is named for its use in cakes called mustaceum. | Mustaceum |
| Nancy howard | Roots are hardy to Philadelphia, Pennsylvania. | Nancy Howard bay |
| Olivaeformis | Fruits are more olive-shaped. | Olive-fruited bay |
| Ovalifolia | Distinctly oval-shaped leaf. | Oval-leaved bay |
| Pallida | Leaves are yellow-green; fruits have reddish, shrubby growth. | |
| Parvifolia | Small leaves; French origin Caribbean. | Small-leaved bay |
| Pedunculata | Flowers are more pedunculate. | Pedunculate-flowered bay |
| Popeye | Narrowly pyramidal tree with shorter internodes and compact for small spaces. | |
| Pride of Provence | Compact growing bay tree with lush dark green shiny foliage; shorter internodes, requiring little or no trimming. | Hedge bay laurel |
| Rotundifolia | Leaves are wide, mostly round. | Round-leaved bay |
| Rubrinervis | Red-veined leaves. | Red-veined bay |
| Salicifolia | Narrow lance-shaped leaves are not as thick as the normal variety, have a lighter green color, shrubby growth (1.8 to 2.4 m); Mostly confused in nursery trade with ‘Angustifolia.’ | Willow-leaf bay |
| Saso’s dwarf | Leaves are thick, dark green; shrubby growth | |
| Sphaerocarpa | Leaves are small, polished, spherical fruits. | Ball-fruited bay |
| Sunspot | Gold-variegated foliage. | Sunspot sweet bay |
| Variegata | gold-striped bay |
| Leaves | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Location | 1,8–Cineole | Sabinene | α-Terpinyl Acetate | α-Pinene | Linalool | Methyl Eugenol | β-Pinene | Eugenol | Camphene | α-Terpineol | Reference |
| Albania | 26.70 | 11.80 | 12.00 | 2.20 | 18.50 | 2.50 | 2.40 | 6.50 | 0.20 | 1.20 | [57] |
| Algeria | 17.6–44.13 | 2.20–9.60 | 7.90–17.33 | 0.90–9.20 | 4.18–12.57 | 5.10–11.0 | 0.80–3.80 | 1.20–3.60 | 0.20–8.91 | 2.58–7.6 | [58,59] |
| Argentina | 37.3–43.8 | 8.4 | 7.9–10.6 | 4.8 | 12.7–19.4 | 3.0–3.6 | 3.5–20.1 | 0.7 | 0.4 | 2.2–2.8 | [60] |
| Brazil | 26.9–37.3 | 10–4−13.7 | 15.3–17.4 | 8.9–10.9 | 0.4–0.6 | 0.71–0.72 | - | 0.39 | 0.44–0.51 | 0.04 | [61] |
| Bulgaria | 41.0 | 8.8 | 14.4 | 2.56 | 4.92 | 6.0 | 2.45 | 1.47 | 0.18 | 3.11 | [62] |
| China | 25.5–43.0 | 2.1–6.8 | 9.5–18.0 | 1.0–5.2 | 4.4–22.7 | 1.7–8.9 | 1.0–4.4 | 1.4–2.4 | 0.2 | 1.5–2.5 | |
| Colombia | 22.0 | 1.5 | 11.1 | 2.9 | 16.4 | 2.9 | 6.1 | 2.0 | 0.1 | 4.9 | |
| Croatia | 45.5 | 5.7 | 9.1 | 2.1 | 8.5 | 10.0 | -- | 2.5 | -- | 1.5 | [63] |
| France | 39.1 | 4.4 | 18.2 | 2.2 | 10.0 | 11.8 | 1.7 | -- | 0.2 | 1.3 | [64] |
| Germany | 23.3 | 5.3 | 9.8 | 4.6 | 2.0 | 3.5 | 4.0 | 2.9 | 0.7 | 3.3 | [65] |
| Greece | 49.60 | 7.80 | 5.25 | 5.96 | 1.50 | 2.10 | 5.12 | 5.60 | 1.21 | 1.90 | [66,67] |
| India | 0.27–3.31 | 0.22 | -- | 1.37–7.68 | 29.08–50.68 | 1.00 | 0.71–1.52 | 63.57 | 0.26–1.14 | 0.39–0.60 | [68] |
| Iran | 25.7–61.0 | 2.33–8.7 | 6.14–15.14 | 2.35–6.59 | 1.40–3.96 | 3.08–5.18 | 1.40–4.6 | 0.4–2.88 | 0.27–10.22 | 0.69–3.88 | [69,70,71,72,73] |
| Israel * | 1698.2–2549.7 | 168.8–277.1 | 255.6–482.3 | 58.6–77.6 | 106.7–151.5 | 36.8–40.2 | 37.2–69.2 | 7.5–11.8 | 4.2–4.5 | 184.5–233.1 | |
| Italy | 22.84–35.70 | 4.30–6.50 | 4.43–14.23 | 2.6–5.72 | 7.08–19.47 | 2.52–16.22 | 2.40–3.46 | 1.6–5.97 | 0.14–0.30 | 2.42–6.44 | [74,75,76] |
| Jordan | 36.80–40.91 | 3.10–6.92 | 5.86–14.6 | 4.60–5.82 | 1.29–2.60 | 1.62–4.20 | 3.60–4.55 | 0.92 | 0.50–0.58 | -- | [17,77] |
| Morocco | 35.62–58.88 | 0.42–6.13 | 0.45–8.96 | 3.72–4.58 | 1.98–9.45 | 1.70–3.93 | 1.92–3.14 | 0.56–1.97 | 0.5–4.87 | 1.56–5.83 | [78,79] |
| Lebanon | 57.05–65.99 | 4.06–9.74 | -- | 2.14–6.03 | 0.51–0.75 | -- | 2.51–4.17 | -- | 0.14–0.17 | 2.90–3.64 | |
| Nepal | 1.64–26.64 | 0.26–0.43 | -- | 2.05–6.70 | 28.97–72.67 | -- | 1.17–3.03 | 0.21–0.23 | 0.49–1.68 | 0.37–3.05 | [52] |
| Portugal | -- | 4.00 | 10.20 | 2.3 | 8.4 | 5.4 | 1.8 | 1.2 | 0.1 | 2.7 | [80] |
| Turkey | 46.61–72.09 | 4.44–14.05 | 4.04–25.70 | 2.19–6.11 | 0.37–1.9 | 0.41–3.39 | 2.58–3.91 | 0.40–1.55 | 0.19–0.67 | 0.95–6.83 | [3,14] |
| Spain | 33.28–43.56 | 2.48–5.80 | 6.0–11.75 | 0.85–11.6 | 6.50–26.70 | 3.12–4.64 | 3.47–4.90 | 1.23–3.00 | 0.7–1.06 | 0.9–4.95 | [57,81] |
| Serbia | 15.54–41.86 | 0.55–9.12 | 5.49–24.74 | 0.12–7.20 | 1.81–16.00 | 5.32–8.67 | 0.16–5.23 | 1.07–6.14 | 0.06–0.83 | 1.65–4.28 | [82,83,84,85] |
| Syria | 58.66–73.70 | 3.56–8.89 | -- | 2.62–3.85 | 0.32–0.97 | -- | 2.66–3.25 | -- | 0.05–0.27 | 1.12–3.50 | [17] |
| Fruit | |||||||||||
| Location | 1,8–Cineole | Sabinene | α–Terpinyl acetate | α–Pinene | (E)-β-ocimene | α–Phellandrene | β-Pinene | β-Elemene | Camphene | Germacrene-A | Reference |
| Bulgaria | 33.33 | 6.30 | 10.30 | 11.01 | 0.72 | 5.18 | 0.28 | 7.45 | 4.33 | [62] | |
| Turkey | 9.50–20.45 | 1.70–6.03 | 1.20–4.88 | 3.3–16.55 | 11.88–28.35 | 10.58–15.87 | 2.1–12.83 | 2.0–4.46 | 0.80–2.08 | 0.80–4.35 | [3,12] |
| Jordan | 29.8 | 4.4 | 1.2 | 10.9 | 3.2 | 9.0 | 8.4 | 6.2 | 1.3 | 0.2 | [77] |
| Lebanon | 17.64–48.01 | 2.93−4.49 | 0.87–2.07 | 7.69–17.96 | 0.57–11.82 | 8.29–17.07 | 3.91–9.51 | - | 1.08–2.61 | - | |
| Iran | 14.4–46.7 | 5.4 | 5.8.5 | 2.8–6.6 | 20.8–22.1 | 4.7 | 5.1–7.3 | 2.1–3.5 | [86] | ||
| Tunisia | 8.1–8.8 | 1.8–2.6 | 3.0–3.8 | 8.0–10.3 | 20.9–23.7 | 4.2–5.8 | 2.6–3.8 | [87] | |||
| Israel * | 800.0–1278.5 | 97.6–103.1 | 312.4–340.1 | 1262.9–1284.6 | 204.7–212.6 | 228.5–242.0 | 411.8–373.1 | 53.2–55.5 | 904.2–1012.9 | ||
| Flower | |||||||||||
| Location | 1,8–Cineole | Sabinene | α–Terpinyl acetate | α–Pinene | (E)-β-ocimene | Linalool | β-Pinene | β-Elemene | Bornyl acetate | E-β-Caryophyllene | Reference |
| Turkey | 8.8 | 1.7 | 1.8 | 5.1 | 2.7 | - | 3.7 | 5.4 | 2.1 | 5.1 | [12] |
| Italy | 7.9–42.8 | 0.5–6.0 | 0.3–12.0 | 0.9–3.8 | 0.1–65.3 | 0.6–14.4 | 0.4–2.9 | 0.1–5.3 | 0.1–0.6 | 0.1–15.4 | [88] |
| Morocco | 45.01 | 3.01 | 0.1 | 3.04 | - | 1.04 | 3.01 | - | 0.25 | 0.42 | [89] |
| Israel * | 1186.4–3526.1 | 328.5–344.1 | 1314.4 | 278.7–452.1 | 1046. | 346.7 | 401.4 | 421.0–498.1 | 1019.7 | 1715.1 | |
| Sample Number | Extraction Method | Features/Treatment | Effect on Volatiles Composition | Reference |
|---|---|---|---|---|
| 1 | Enzyme assisted extraction | Pre-treatment with cellulase, hemicellulase, xylanase for 1 h at 40 °C | Increase in 1,8-cineole, methyl eugenol, terpinen-4-ol, α terpineol, and caryophyllene oxide than hydrodistillation | [94] |
| 2 | Simultaneous distillation extraction (SDE) | Extraction for 2 h in dichloromethane using microscale simultaneous distillation apparatus | More hydrocarbon monoterpenes (15.7%) as with from SPME (3.6%) | [81] |
| 3 | Solid-phase microextraction (SPME) | Adsorption in headspace using dimethylsiloxane fiber (100 µm) at 60 °C for 30 min | Oxygen terpenes (95.7%) as compared with SDE (83.4%) | [81] |
| Sample 3 mg instead of 50–200 mg | Identified 98 different volatiles | [76] | ||
| 4 | Supercritical carbon dioxide (SFE) | Pressure, 100 bar; temperature, 40 °C; and CO2 flow, φ 0.3 kg/h; time, 1.4 h | Two times less monoterpene hydrocarbons and oxygenated monoterpenes (43.89%) in comparison to HD (98.4%). | [84] |
| Pressure, 100/250 bar; temperature, 40/50 °C; and CO2 flow, φ 1.629 dm3/min; time, 3 h | 100 bar/40 °C and 250 bar/40 °C had the highest amount of 1, 8-cineole, but at 100 bar/60 °C, α-terpineol acetate was dominant. | [82] | ||
| Pressure, 90 bar; temperature, 50 °C; and CO2 flow, φ 1.0 kg/h. | Monoterpenes were extracted in shorter times than sesquiterpenes | [74] | ||
| Pressure, 80 bar; temperature, 40 °C and 50 °C; and CO2 flow, φ 1 mL/min; time, 20 min. | A decrease in monoterpene hydrocarbons (6.8–5.1%) in comparison to hydro–(19.5%) and steam (16.5%) distilled oils | [95] | ||
| 5 | Solvent-Free Microwave Extraction (SFME) | Power, 850 W for 30 min | 1,8-cineole, linalool, eugenol and Methyl eugenol content increased | [96] |
| Power, 300, 600, and 900 W for 20 min | Oxide volatile organic compound reduced in comparison to HD. The highest 1,8-cineole (72%) content at 300 W | [96] | ||
| Power, 622 and 249 W for 1 h | No significant differences in volatiles by SFME and HD but 55–60% time was reduced by SFME | [96] | ||
| Power, 200, 300 W and pulsed microwaves system for 1 h | Higher amounts of oxygenated compounds and lower amounts of monoterpenes | [76] | ||
| 6 | Optimum ohmic heating assisted hydrodistillation (OAHD) | 120 min, 8.53 V/cm, and 40 g | EO yield of OAHD was found to be higher when compared with HD methods | [97] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paparella, A.; Nawade, B.; Shaltiel-Harpaz, L.; Ibdah, M. A Review of the Botany, Volatile Composition, Biochemical and Molecular Aspects, and Traditional Uses of Laurus nobilis. Plants 2022, 11, 1209. https://doi.org/10.3390/plants11091209
Paparella A, Nawade B, Shaltiel-Harpaz L, Ibdah M. A Review of the Botany, Volatile Composition, Biochemical and Molecular Aspects, and Traditional Uses of Laurus nobilis. Plants. 2022; 11(9):1209. https://doi.org/10.3390/plants11091209
Chicago/Turabian StylePaparella, Antonello, Bhagwat Nawade, Liora Shaltiel-Harpaz, and Mwafaq Ibdah. 2022. "A Review of the Botany, Volatile Composition, Biochemical and Molecular Aspects, and Traditional Uses of Laurus nobilis" Plants 11, no. 9: 1209. https://doi.org/10.3390/plants11091209